真核基因表达调控优质PPT.ppt
《真核基因表达调控优质PPT.ppt》由会员分享,可在线阅读,更多相关《真核基因表达调控优质PPT.ppt(147页珍藏版)》请在冰豆网上搜索。
表达水平降低eg:
-半乳糖苷酶意义:
适应环境,基因表达的时空特异性,1、时间特异性(temporalspecificity),2、空间特异性(spatialspecificity),在个体生长全过程,某种基因产物在个体按不同组织空间顺序出现,称之为基因表达的空间特异性,又称细胞或组织特异性(cellortissuespecificity)。
真核基因的表达调控的特点:
原核细胞环境因素对调控起决定性的作用。
群体中每一个细胞对环境变化的反应是直接的和一致的。
真核细胞基因表达调控最明显的特征是在特定时间,特定的细胞中特定的基因被激活,实现“预定”的、有序的、不可逆转的分化、发育,并使生物的组织和器官保持正常功能。
这是生命活动规律决定的,环境因素在其中作用不大。
真核生物基因调控可分为两大类:
第一类是瞬时调控或称可逆性调控,它相当于原核细胞对环境条件变化所做出的反应,包括某种底物或激素水平升降,或细胞周期不同阶段酶活性的调节;
第二类是发育调控或称不可逆调控,是真核基因调控的精髓部分,决定了真核细胞生长、分化、发育的进程。
根据基因调控在同一事件中发生的先后次序又可分为:
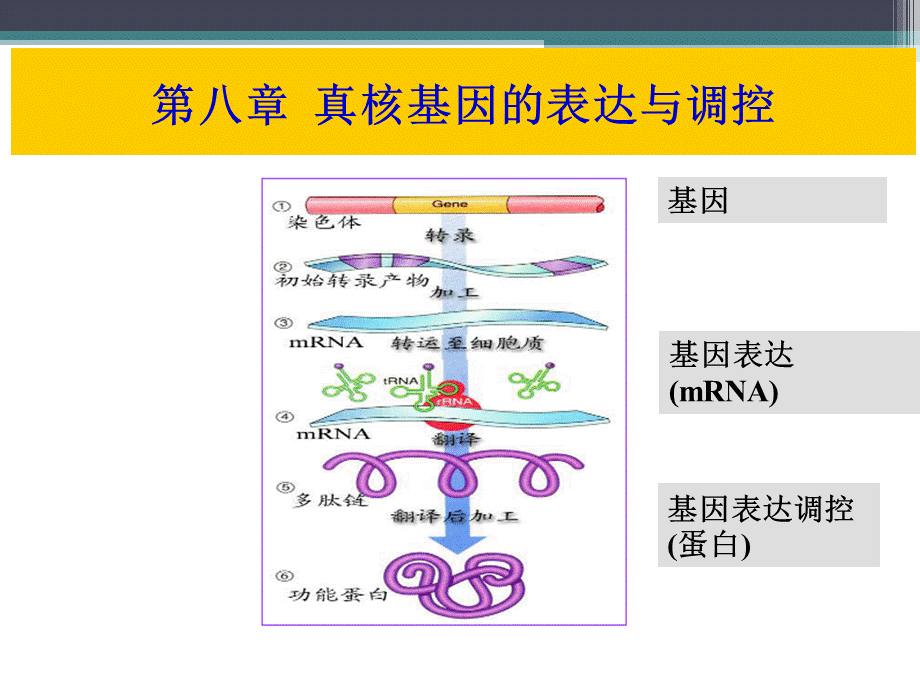
DNA水平调控转录水平调控转录后水平调控翻译水平调控蛋白质加工水平的调控,2.真核基因转录水平表达调控,一、真核基因转录
(一)真核基因结构,“基因”的分子生物学定义:
产生一条多肽链或功能RNA所必需的全部核苷酸序列。
真核基因调控主要在转录水平上进行,受大量特定的顺式作用元件(cis-actingelement)和反式作用因子(transactingfactor)调控。
真核生物的转录调控大多数是通过顺式作用元件和反式作用因子复杂的相互作用来实现的。
(2)增强子:
指能提高转录起始频率的DNA序列。
SV40的转录单元上发现,转录起始位点上游约200bp处有两段长72bp的正向重复序列。
增强子特点:
(P89),增强子对转录的影响增强子的功能受DNA双螺旋空间构象的影响。
增强子可能有如下3种作用机制:
影响模板附近DNA双螺旋结构,导致DNA双螺旋弯折或在反式因子的参与下,以蛋白质之间的相互作用为媒介形成增强子与启动子之间“成环”连接,活化基因转录。
将模板固定在细胞核内特定位置,如连接在核基质上,有利于DNA拓扑异构酶改变DNA双螺旋结构的张力,促进RNA聚合酶在DNA链上的结合和滑动。
增强子区可以作为反式作用因子或RNA聚合酶进入染色质结构的“入口”。
沉默子(silencer),沉默子是可降低基因启动子转录活性的一段DNA顺式元件。
与增强子作用相反。
沉默子的DNA序列被调控蛋白结合后阻断了转录起始复合物的形成或活化,使基因表达活性关闭,绝缘子(insulator),绝缘子(insulator)长约几百个核苷酸对,是通常位于启动子同正调控元件(增强子)或负调控因子(为异染色质)之间的一种调控序列。
绝缘子本身对基因的表达既没有正效应,也没有负效应,其作用只是不让其他调控元件对基因的活化效应或失活效应发生作用。
绝缘子的作用是有方向性的,,反式作用因子,1、定义:
能直接或间接地识别或结合在各类顺式作用元件核心序列上,参与调控靶基因转录效率的蛋白质。
TFD(TATA)、CTF(CAAT)、SP1(GGGCGG)、HSF(热激蛋白启动区),转录调节因子,识别增强子、沉默子等。
通过DNA-蛋白质相互作用,,基本转录因子,识别启动子,通过DNA-蛋白质相互作用。
共调节因子,通过蛋白质-蛋白质相互作用,协调调控,1.分类,转录因子:
激活因子、阻遏因子共调节因子:
共激活因子、共阻遏因子,2、结构,DNA结合结构域,转录活化结构域,结构域,结构域是指蛋白质亚基结构中明显分开的紧密球状结构区域,又称为辖区。
酵母双杂交由Fields在1989年提出.他的产生是基于对真核细胞转录因子特别是酵母转录因子GAL4性质的研究.GAL4包括两个彼此分离的但功能必需的结构域.位于N端1-174位氨基酸残基区段的DNA结合域(DNAbindingdomain,DNA-BD)和位于C端768-881位氨基酸残基区段的转录激活域(Activationdomain,AD).DNA-BD能够识别位于GAL4效应基因(GAL4-responsivegene)的上游激活序列(Upstreamactivatingsequence,UAS),并与之结合.而AD则是通过与转录机构中的其他成分之间的结合作用,以启动UAS下游的基因进行转录.DNA-BD和AD单独分别作用并不能激活转录反应,但是当二者在空间上充分接近时,则呈现完整的GAL4转录因子活性并可激活UAS下游启动子,使启动子下游基因得到转录。
Fields建立了一个双杂交系统,DB与X蛋白融合,AD与Y蛋白融合,如果X、Y之间形成蛋白-蛋白复合物,使GAL4两个结构域重新构成,启动特异基因序列的转录,该实验的结果表明由X和Y相互作用使得AD和BD在空间上接近,激活了报告基因LacZ的转录.一般地,将DB-X的融合蛋白称作诱饵(bait),X往往是已知蛋白,AD-Y称作猎物(prey),能显示诱饵和猎物相互作用的基因称报告基因,通过对报告基因的检测,反过来可判断诱饵和猎物之间是否存在相互作用.,DNA结合结构域:
模体螺旋-转折-螺旋(Helix-turn-helix,H-T-H)锌指结构(zincfinger)碱性-亮氨酸拉链(basic-leucinezipper)碱性-螺旋-环-螺旋(basichelix/loop/helix,bHLH),1、螺旋-转折-螺旋(HTH),这类蛋白质分子中有至少两个螺旋,中间由短侧链氨基酸残基形成“转折”,蛋白质能识别DNA,主要是靠着蛋白质氨基酸侧链与DNA的碱基之间形成氢键以及疏水性作用来完成;
识别时不必打开双螺旋。
红色部分为识别螺旋,蓝色部分则起帮助识别螺旋定位到DNA大沟上的作用。
2、锌指结构,配位键,2-9个,由大约30个氨基酸残基的肽段与锌螯合形成的指形结构。
锌以四个配位键与肽链的Cys或His残基结合,指形突起的肽段含12-13个氨基酸残基,嵌入DNA的大沟中,由指形突起或其附近的某些氨基酸侧链与DNA的碱基结合而实现蛋白质与DNA的结合。
TFIIIA,Cys2/His2锌指,见于SP1,TFA等,Cys2/Cys2锌指,见于甾体激素受体,344aa,N端与DNA结合9个锌指,每个30aa与5srRNA基因内启动子(50bp)结合,TFIIIA,转录因子SP1(GC盒)、连续的3个锌指重复结构。
3、碱性-亮氨酸拉链,二聚体亮氨酸之间相互作用形成二聚体,形成“拉链”。
肽链氨基端2030个富含碱性氨基酸结构域与DNA结合。
这类蛋白质的DNA结合结构域实际是以碱性区和亮氨酸拉链结构域整体作为基础的。
即bZIP结构。
蛋白中每隔6个氨基酸就有一个亮氨酸残基,导致第7个亮氨酸残基都在螺旋的同一方向出现。
定义:
出现在DNA结合蛋白质和其它蛋白质中的一种结构基元(motif)。
当来自同一个或不同多肽链的两个-螺旋的疏水面(常常含有亮氨酸残基)相互作用形成一个圈对圈的二聚体时就形成了亮氨酸拉链。
4、碱性-螺旋-环-螺旋,在免疫球蛋白轻链基因的增强子结合蛋白E12与E47中,羧基端100200个氨基酸残基可形成两个两性螺旋,被非螺旋的环状结构所隔开,蛋白质的氨基端则是碱性区,其DNA结合特性与亮氨酸拉链类蛋白相似。
bHLH类蛋白只有形成同源或异源二聚体时,才具有足够的DNA结合能力。
当这类异源二聚体中的一方不含有碱性区(如Id或E12蛋白)时,该二聚体明显缺乏对靶DNA的亲和力。
亮氨酸拉链(leucinezipper)螺旋-环-螺旋(H-L-H)结构,转录活化结构域,转录激活结构域(transcriptionactivationdomains)一般由30-100个氨基酸残基组成,负责结合并激活别的转录调控蛋白。
如GAL4分子中有2个这种结构域,分别位于多肽链的第147-196位和第768-881位.,在不同的转录活化域有下列特征性结构:
带负电荷的螺旋结构。
哺乳动物细胞中糖皮质激素受体的两个转录活化域都有酸性的螺旋结构,它们可能与TFIID复合物中某个通用因子或RNA聚合酶II本身结合,具有稳定转录起始复合物的作用。
富含谷氨酰胺的结构。
SP1是启动子GC盒的结合蛋白,共有4个参与转录活化的区域,其中最强的转录活化域含25%左右的谷氨酰胺富含脯氨酸的结构。
CTF-NF1因子羧基端富含脯氨酸(达20%30%)。
在Oct2、Jun、AP2、SRF等哺乳动物因子中也有富含脯氨酸的结构域。
3.染色体及DNA水平的调控,异染色质化p327,异染色质是包装成20-30nm,不具有转录活性的染色质组成性异染色质(constitutiveheterochromatin)是指在各种细胞中,在整个细胞周期内都处于凝聚状态的染色质,如着丝粒,端粒,核仁形成区等。
兼性异染色质(facultativeheterochromatin)指在某些特定的细胞中,或在一定的发育时期和生理条件下凝聚,由常染色质变成异染色质,这本身也是真核生物的一种表达调控的途经。
莱昂假说(Lyonhypothesis),巴氏小体是一个失活的X染色体,失活的过程就称为莱昂化(lyonization);
在哺乳动物中,雌性个体细胞中的两个X染色体中有一个X染色体在受精后的第16天(受精卵增殖到5000-6000,植入子宫壁时)失活;
两条X染色体中哪一条失活是随机的;
X染色体失活后,细胞继续分裂形成的克隆中,此条染色体都是失活的;
生殖细胞形成时失活的X染色体可得到恢复。
活性染色质对DNase的敏感性p316,可以用DNase来鉴别基因的活性,组蛋白对基因活性的影响,组蛋白的乙酰化和去乙酰化控制染色质活性p323-324,组蛋白乙酰化是活性染色质的标志之一,去乙酰化和基因活性的阻遏有关,组蛋白的乙酰化及去乙酰化对基因表达的影响组蛋白乙酰化的状态与基因表达有关。
组蛋白N端“尾巴”上赖氨酸残基的乙酰化中和了组蛋白尾巴的正电荷,降低了它与DNA的亲和性,导致核小体构象发生有利于转录调节蛋白与染色质相结合的变化,从和提高了基因转录的活性。
相反,组蛋白去乙酰化与基因活性的阻遏有关。
组蛋白乙酰基转移酶和去乙酰化酶只能有选择地影响一部分基因的转录。
组蛋白的甲基化与非活性染色质有关,基因丢失:
在细胞分化过程中,可以通过丢失掉某些基因而去除这些基因的活性。
某些原生动物、线虫、昆虫和甲壳类动物在个体发育中,许多体细胞常常丢失掉整条或部分的染色体,只有将来分化产生生殖细胞的那些细胞一直保留着整套的染色体。
基因扩增:
基因扩增是指某些基因的拷贝数专一性增大的现象,它使得细胞在短期内产生大量的基因产物以满足生长发育的需要,是基因活性调控的一种方式。
如非洲爪蟾体细胞中rDNA的基因拷贝数约有500个,而在卵母细胞中的拷贝数约为200万个,是因发育需要出现的基因扩增现象。
基因组拷贝数增加,即多倍性,在植物中是非常普遍的现象。
基因组拷贝数增加使可供遗传重组的物质增多,这可能构成了加速基因进化、基因组重组和最终物种形成的一种方式。
发育或系统发生中的倍性增加在植物中普遍存在,将一个基因从远离启动子的地方移到距它很近的位点从而启动转录,这种方式被称为基因重排。
通过基因重排调节基因活性的典型例子是免疫球蛋白结构基因的表达。
基因重排:
轻链(L)和重链(H)可变区(V)和恒定区(C)多样区(D)铰链区(J),免疫球蛋白分子的基本结构,单体、双体和五聚体,基因重排与变换免疫球蛋白重链基因片段重排与组织特异性表达,基因重排,DNA水平上的调控p319,DNA的甲基化,5-甲基胞嘧啶(5-mC)N6-甲基腺嘌呤(N6-mA)7-甲基鸟嘌呤(7-mG),在真核生物中,5-甲基胞嘧啶主要出现在CpG序列中,CpG二核苷酸通常成串出现在DNA上,CpG岛,真核生物细胞内存在两种甲基化酶活性:
日常型甲基转移酶,从头合成型甲基转移酶,日常型甲基转移酶,DNA甲基化抑制基因转录的机理,DNA甲基化导致某些区域DNA构象变化,从而影响了蛋白质与DNA的相互作用,抑制了转录因子与启动区DNA的结合效率。
DNA甲基化与染色体失活雌性哺乳动物体细胞的两条X染色体中会有一条发生随机失活这是一种基因剂量补偿的机制。
X染色体的Xq13区段(巴氏小体浓缩部位)有一个X失活中心(X-inactioncenter,Xic),X-失活从Xic区段开始启动,然后扩展到整条染色体。
失活基因Xist去甲基化,表达,X染色体的其他位点甲基化,基因不表达,其它水平上的基因调控,1.RNA的加工成熟各种基因的转录产物都是RNA,无论是rRNA、tRNA还是mRNA,初级转录的产物只有经过加工,才能成为有生物功能的活性分子。
1.1rRNA和tRNA的加工成熟rRNA加工有两个内容,一个是分子内的切割,另一个是化学修饰。
rRNA的化学修饰主要是核糖甲基化。
tRNA基因转录时也可能先生成前体tRNA,tRNA基因的初级转录产物在进入细胞质后,首先经过核苷的修饰,生成4.5S前体tRNA,再剪接成为成熟tRNA(4S)。
1.2mRNA的加工成熟编码蛋白质的基因转录产生mRNA。
这类基因产物在转录后要进行一系列的加工变化,才能成为成熟的有生物功能的mRNA。
这些加工主要包括在mRNA的5末端加帽子,在其3末端加上poly(A),进行RNA的剪接以及核苷酸的甲基化修饰等。
由于mRNA的这些结构与它作为蛋白质合成模板的功能有密切关系,所以是基因表达的重要调控环节。
编码蛋白质的基因转录时首先生成前体pre-mRNA(或称核不均一RNA,hnRNA),然后再加工剪接为成熟mRNA。
1.3真核生物基因转录后加工的多样性真核生物的基因可以按其转录方式分为两大类:
简单转录单位复杂转录单位。
这两种转录方式虽然最终都产生蛋白质,但它们的转录后加工方式是不同的。
简单转录单位。
这类基因只编码产生一个多肽,其原始转录产物有时需要加工,有时则不需要加工。
有3种不同形式:
第一种简单转录单位,如组蛋白基因,它们没有内含子,因此不存在转录后加工问题,其mRNA3末端没有poly(A),但有一个保守的回文序列作为转录终止信号。
第二种简单转录单位包括腺病毒蛋白IX、-干扰素和许多酵母蛋白质基因,它们没有内含子,所编码的mRNA不需要剪接,但需要加poly(A)。
第三种简单转录单位包括和-珠蛋白基因及许多细胞蛋白基因,这些基因虽然都有内含子,需要进行转录后加工剪接,还要加poly(A),但它们只产生一个有功能的mRNA,所以仍然是简单转录单位。
简单转录单位:
只编码产生一个多肽,复杂转录单位。
含有复杂转录单位的主要是一些编码组织和发育特异性蛋白质的基因,它们除了含有数量不等的内含子以外,其原始转录产物能通过多种不同方式加工成两个或两个以上的mRNA。
也有三种情况:
利用多个5端转录起始位点或剪接位点产生不同的蛋白质利用多个加poly(A)位点和不同的剪接方式产生不同的蛋白质。
这类基因调控点不在5末端和内含子的不同剪接。
而在于有两个或多个加poly(A)位点,因此可通过不同的剪接方式得到不同的蛋白质。
虽无剪接,但有多个转录起始位点或加polyA位点的基因,前mRNA不同的剪接方式造成了不同组织中不同的降钙素样蛋白,复杂转录单位:
产生至少2种mRNA,1.4mRNA有效性的调控真核生物能否长时间、及时地利用成熟的mRNA分子翻译出蛋白质以供生长、发育的需要,是与mRNA的稳定性以及屏蔽状态的解除密切相关的。
原核生物mRNA的半衰期很短,平均大约3min。
高等真核生物迅速生长的细胞中mRNA的半衰期平均约为3h。
在高度分化的终端细胞中许多mRNA极其稳定,有的寿命长达几天或十几天,加上强启动子的转录,使一些终端细胞特有的蛋白质合成达到惊人的水平。
例如,家蚕丝心蛋白基因具有很强的启动子,几天内即可转录出105个丝心蛋白mRNA,而它的寿命长达4天,每个mRNA分子能重复翻译出105个丝心蛋白,所以4天内可产生1010个丝心蛋白,说明mRNA寿命的延长是mRNA有效性的一个重要因素。
mRNA有效性的调控原核生物mRNA的半衰期很短,平均大约3min。
mRNA寿命的延长是mRNA有效性的一个重要因素。
翻译水平的调控在蛋白质生物合成的起始反应中主要涉及到细胞中的4种装置,这就是:
核糖体,它是蛋白质生物合成的场所蛋白质合成的模板mRNA,它是传递基因信息的媒介可溶性蛋白因子,这是蛋白质生物合成起始物形成所必需的因子tRNA,它是氨基酸的携带者。
只有这些装置和谐统一才能完成蛋白质的生物合成。
1.真核生物mRNA的“扫描模式”与蛋白质合成的起始AUG的前和后序列特征为A/GNNAUGG,这使得核糖体滑行到mRNA的第一个AUG能准确识别并起始翻译。
2.mRNA5末端帽子结构的识别与蛋白质合成,3.mRNA的稳定性与基因表达调控4.可溶性蛋白因子的修饰与翻译起始调控许多可溶性蛋白因子(起始因子),对蛋白质合成的起始有着重要的作用,对这些因子的修饰会影响翻译起始。
在蛋白质生物合成的过程中,特别是在起始反应中,mRNA的“可翻译性”是起决定作用的,其5末端的帽子结构、二级结构、与rRNA的互补性以及起始密码附近的核苷酸序列都是蛋白质生物合成的信号系统。
小结,真核生物的基因组真核生物基因表达调控的特点和种类真核生物DNA水平上的基因表达调控真核生物转录水平上的基因表达调控真核基因转录后水平上的调控,顺式作用元件和反式作用因子,RNA的加工成熟、翻译水平的调控,DNA结合结构域常见的有、DNA水平上的调控包括、等方式。
其中免疫球蛋白结构基因是的典型例子,非洲爪蟾的卵母细胞中rRNA基因的变化是的例子。
真核生物基因调控主要在水平进行的,是通过和复杂的相互作用来实现的。
基因转录的有启动子中的、。
RNA的剪接分为和。
基因表达根据是否受时期、部位、环境影响可以分为和,1.目前认为基因表达调控的主要环节是()A.基因活化B.转录起始C.转录后加工D.翻译起始2.处于活跃转录状态的真核基因对DNaseI()A.高度敏感B.中度敏感C.低度敏感D.不敏感3.关于反式作用因子下列说法正确的是()A.是具有激活功能的调节蛋白B.通过蛋白质-蛋白质相互作用,对翻译进行调控C.通过RNA-蛋白质相互作用,对转录进行调控D.通过DNA-蛋白质或者蛋白质-蛋白质相互作用,对转录进行调控4.在RNA聚合酶II的基本转录因子中,能直接识别、结合TATAbox的是()A.TFIIAB.TFIIBC.TFIIDD.TFIIE5.下列不属于反式作用因子的DNA结合结构域的是()A.bZIPB.HTHC.碱性-螺旋结构D.酸性-螺旋结构,1.所有高等真核生物的启动子中都含有TATAbox结构。
()2.高等真核生物的大部分DNA是不编码蛋白质的。
()3.大多数的管家基因编码低丰度的mRNA()4.人类基因组含有35000个基因,编码35000个蛋白质()5.真核生物基因的mRNA是多顺反子()6.DNA甲基化导致DNA构象变化,从而抑制转录因子与启动区的结合效率()7.Xist基因中含有大量终止密码子,不编码蛋白质()8.有活性的X染色体上的Xist位点都没有甲基化,而失活的X染色体上的Xist位点高度甲基化(9.CpG岛指该二核苷酸序列成串出现的这段DNA序列()10.免疫球蛋白结构基因是通过基因扩增来调节基因活性的。
),1.一个基因的内含子能够作为另一个基因的外显子。
()2.cDNA分子是由mRNA前体转录来的,因此也有内含子序列()3.DNaseI超敏感位点的产生是染色质结构规律变化的结果。
()4.核心启动子只能确定转录起始点产生基础水平的转录。
()5.转录因子具有独立的DNA结合结构域和转录激活结构域。
()6.真核生物基因组中的重复序列都是不翻译的内含子。
()7.核不均一RNA是mRNA和rRNA的前体而不是tRNA的前体()8.所有真核生物的基因都要经过加尾和内含子的剪接过程,才能翻译成蛋白质。
()9.原核和真核基因中都含有内含子。
()10.锌指结构与DNA的大沟结合。
(),1.增强子和启动子都是顺式作用元件()2.真核生物基因的“加帽”反应在细胞核内完成。
()3.bZIP和bHLH蛋白都是形成二聚体。
(T)4.每个转录因子结合位点被单个转录因子识别。
()5.一个增强子只对一个基因表现增强效应,即具有基因专一性。
()6.含简单转录单位的基因只编码一个多肽。
()7.选择性剪接能使同一基因在不同的组织中特异性表达。
()8.只有活性染色质转录的基因对DNaseI敏感。
()9.DNA甲基化提高了甲基化位点突变的频率。
()10.乙酰化的组蛋白抑制了核小体的浓缩,使转录因子更容易与DNA接触,有利于提高基因的转录活性。
(),细胞是生命活动的基本单位。
细胞通过DNA的复制和细胞分裂将本身所固有的遗传信息由亲代传至子代,实现增殖繁衍。
它们还不断地“感知”环境变化,并对其作出特定的应答。
细胞应答可以分为3个阶段:
外界信息的“感知”,即由细胞膜到细胞核内的信息传递,染色质水平上的基因活性调控,特定基因的表达,即从DNARNA蛋白质的遗传信息传递过程。
第五节蛋白质磷酸化对基因转录的调控,蛋白质的磷酸化与去磷酸化过程是生物体内普遍存在的信息传导调节方式,几乎涉及所有的生理及病理过程,如糖代谢、光合作用、细胞的生长发育、神经递质的合成与释放甚至癌变等等。
真核细胞主要跨膜信号传导途径,细胞表面受体与配体分子的高亲和力特异性结合,能诱导受体蛋白构象变化,使胞外信号顺利通过质膜进入细胞内,或使受体发生寡聚化而被激活。
受体分子活化细胞功能的途径主要有两条:
一是受体本身或受体结合蛋白具有内源酪氨酸激酶活性,胞内信号通过酪氨酸激酶途径得到传递;
二是配体与细胞表面受体结合,通过G蛋白介异的效应系统产生介质,活化丝氨酸/苏氨
 升级会员
升级会员 