50个肿瘤核心基因Word格式文档下载.docx
《50个肿瘤核心基因Word格式文档下载.docx》由会员分享,可在线阅读,更多相关《50个肿瘤核心基因Word格式文档下载.docx(43页珍藏版)》请在冰豆网上搜索。
BRAF
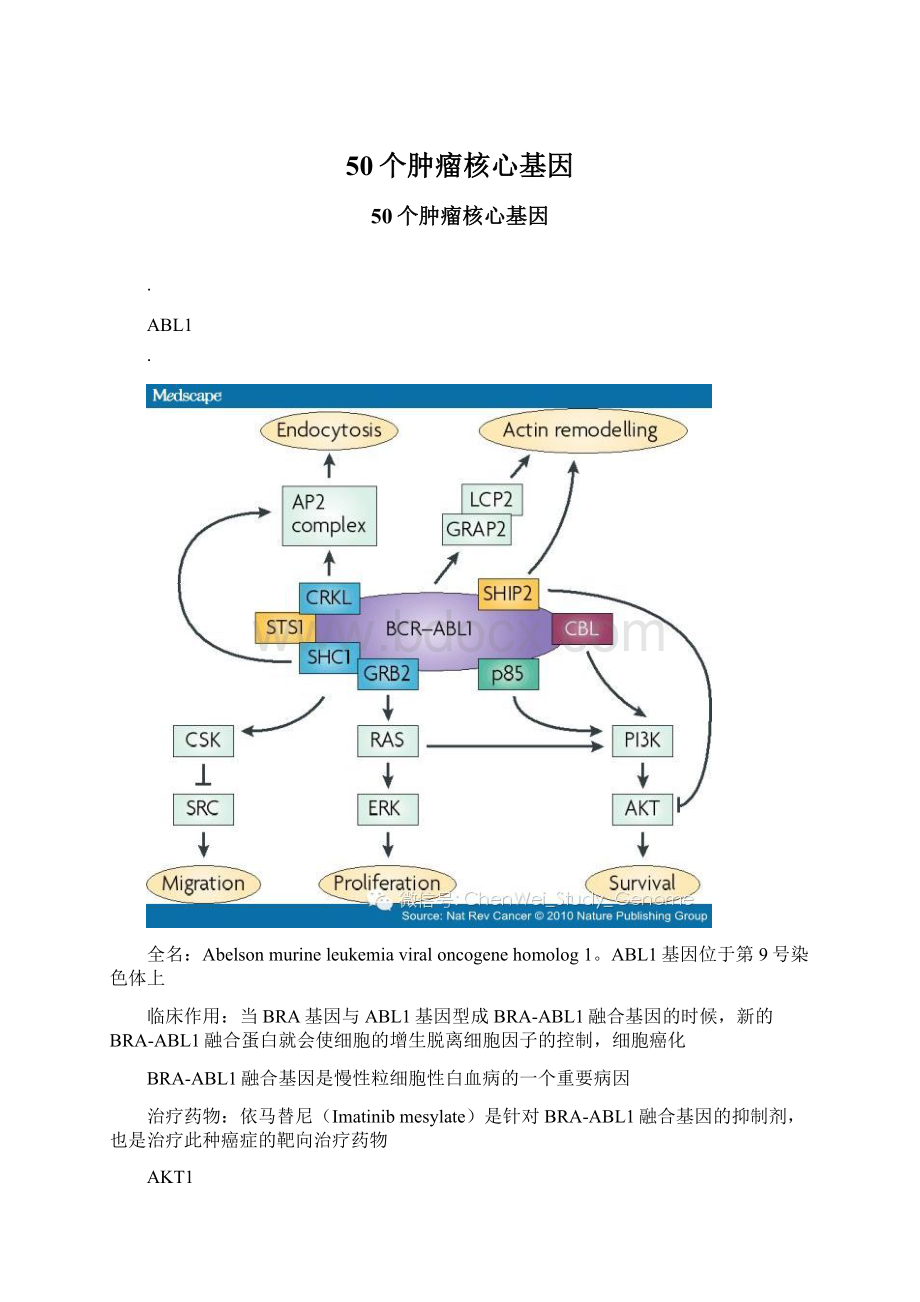
BRAF基因编码B-raf蛋白,是一个激酶,位于7号染色体上
BRAF蛋白激活后导致MEK/ERK的激活,使细胞不进入凋亡程序
BRAF蛋白的完全活化需要T598和S601两个位点的磷酸化。
但是如果BRAF发生V600E突变,那么无需T598和S601两个位点的磷酸化,BRAF就已获得持续的活性,并刺激细胞进入分裂周期
抑制剂,维罗非尼(Vemurafenib)、和索拉菲尼(Sorafenib)是获FDA批准的BRAFV600E突变肿瘤的靶向药物
CDH1
Cadherin-1,位于16号染色体
CDH1是一种依赖于钙离子的细胞粘合蛋白,是一种抑癌基因。
CDH1与癌细胞的浸润、癌细胞转移相关
当CDH1基因发生突变失活时,细胞更容易突破基底膜、侵入到周围的组织中去
CDKN2A
cyclin-dependentkinaseinhibitor2A,又名P16基因,位于9号染色体
CDKN2A是一种抑癌基因,其蛋白可以抑制或延缓细胞从G1期转到S期
CDKN2A的突变与缺失与多种癌症的发生相关
CSF1R
Colonystimulatingfactor1receptor,位于5号染色体
CSF1R编码一个跨膜蛋白,这个蛋白是colonystimulatingfactor1(CSF1)的受体。
CSF1能够促进巨噬细胞的分裂、增殖
CSF1R和慢性骨髓单核细胞性白血病、以及M4急性成髓细胞白血病相关。
CSF1R和CSF1都和乳腺癌相关
EGFR
epidermalgrowthfactorreceptor,又名HER1。
位于7号染色体上。
是重要的细胞生长的细胞膜表面受体。
EGFR是表皮细胞生长的重要调控蛋白,EGFR下游的信号转导通路主要有两条:
一条是Ras/Raf/MEK/ERK-MAPK通路,而另一条是PI3K/Akt/mTOR通路。
多种癌症都与EGFR的过量表达有关系。
EGFR的mRNA表达水平高低,对EGFR靶向治疗效果,会有显著差异。
mRNA表达水平高的非小细胞肺癌患者,往往经针对EGFR的靶向治疗后有良好效果。
而mRNA表达水平低的非小细胞肺癌患都,经EGFR的靶向治疗,往往没有太好的效果。
与治疗药物的关系:
针对EGFR的小分子药物有吉非替尼(gefitinib、iressa)、厄洛替尼(erlotinib、tarceva)和拉帕非尼(lapatinib、tykerb)等。
针对EGFR的单抗药物有西妥昔单抗(cetuximab、erbitux)、帕尼单抗(panitumumab、vectibix)、尼妥珠单抗(nimotuzumab、泰欣生)。
经临床实践证明,上述药物都对多种肿瘤有明显的抑制作用。
基因检测:
各种基因突变,有些会导致耐药、另一些会导致药物敏度。
厦门艾德、上海透景等公司都开发了一系列的突变检测试剂盒,来检测EGFR的各种可能的突变(目前是检测29种突变)。
ERBB2(也称HER2)
Receptortyrosine-proteinkinaseerbB-2,又称HER2,humanepidermalgrowthfactorreceptor2。
位于第17号染色体的长臂。
ERBB2参与MAPK、PI3K/Akt、PKC、STAT等一系列的信号通路。
乳腺癌中30%有ERBB2的突变。
赫赛汀(Herceptin,曲妥珠单抗)是对针对ERBB2的靶向药物,另外帕妥珠单抗(Pertuzumab)也是针对ERBB2的药物。
赫赛汀十分昂贵,而且有心脏毒性,所以用赫赛汀之前应做ERBB2的基因检测,ERBB2基因表达升高的、拷贝数增加的病人,赫赛汀的治疗效果会更好。
erbB4
Receptortyrosine-proteinkinaseerbB-4,又名:
HER4。
位于2号染色体上
和其它的erbB基因一样,erbB4也是一个膜受体蛋白,也同样是酪氨酸激酶
erbB4与多种肿瘤相关尤其与乳腺癌相关
EZH2
Histone-lysineN-methyltransferase,位于7号染色体上
把3个甲基化基团加到组蛋白3(Histone3)的第27号赖氨酸基团上,这会导致染色体的收紧。
与肿瘤的相关性:
EZH2的过量表达会导致多种原发的癌症,原因是它过量表达后,会抑制多种抑癌基因的表达。
FBXW7
F-box/WDrepeat-containingprotein7。
位于4号染色体。
FBXW7直接与细胞周期蛋白E(cyclinE)作用,并很可以导向细胞周期蛋白E参与泛素介导的降解作用。
在卵巢癌、乳腺癌和结直肠癌发现有FBXW7的突变。
FGFR1
▪
Fibroblastgrowthfactorreceptor1。
是成纤维细胞生长因子家族的一员。
这是一个膜受体蛋白,也是一个酪氨酸激酶。
与肺鳞癌相关。
FGFR2
Fibroblastgrowthfactorreceptor2。
它是一个酪氨酸激酶。
一个第2内含子的点突变与一种高乳腺癌风险相关。
FGFR3
Fibroblastgrowthfactorreceptor3。
缺少FGFR3与膀胱癌相关
FLT3
Fms-liketyrosinekinase3,又名:
Clusterofdifferentiationantigen135(CD135).
它是一个三类受体酪氨酸激酶
FLT3的串联持拷贝数增加是急性髓性白血病中最常见的突变,而且有这种突变也意味着预后不好
相关药物:
有发现索拉非尼对FLT3阳性的急性髓性白血病有良好的疗效
GNA11
Guaninenucleotide-bindingproteinsubunitalpha-11。
又名:
Gqα亚基。
位于第9号染色体上。
GNA11是一种G蛋白其功能是激活磷酯酶C(phospholipaseC),进尔参与一系列的信号通路。
GNAS
全名:
G-proteinalphasubunit。
位于20号染色体上。
与脑垂体瘤相关
GNAQ
Guaninenucleotide-bindingproteinG(q)subunitalpha。
HNF1A
hepatocytenuclearfactor1homeoboxA。
位于第12号染色体。
这是一个转录因子,与几种肝脏特异的基因表达调控有关。
HRAS
GTPaseHRas,又名:
transformingproteinp21。
位于第11号染色体。
这是一个小的G蛋白,也是smallGTPases的一员。
一旦结合到GTP上,HRaf就会激活c-Raf之类的Rafkinase,并进一步激活MAPK/ERK通路。
HRAS与膀胱癌有密切的关系HRAS若发生G12V、G12S突变,则其活性就永久激活HRAS的突变与甲状腺癌、肾癌也有关系
IDH1
Isocitratedehydrogenase1(NADP+,异柠檬酸脱氢酶)。
该基因位于2号染色体上。
异柠檬酸脱氢酶催化异柠檬酸转变成戊邻酮二酸与肿瘤的相关性:
IDH1的突变会导致干骺端软骨瘤病,并且IDH1与胶质母细胞瘤相关.
JAK2
Januskinase2。
该基因位于9号染色体。
这是一种不依赖于受体的酪氨酸激酶。
在白血病人中发现在TEL-JAK2融合基因。
JAK3
Januskinase3。
该基因位于第19号染色体
JAK3是一个酪氨酸激酶,主要在造血细胞中表达,如NK细胞、T细胞和B细胞,这一点与同基因家族的JAK1、JAK2在各种细胞中的广泛表达不一样。
JAK3主要在免疫细胞中表达,主要是在被白介素激活后发挥其酪氨酸激酶的功能而起作用。
JAK1、JAK2、JAK3的活化突变,都已被确认是血液性的癌症的病因。
药物:
有一种编号为CP-690550(Tofacitinib,星熠艾克)的JAK3抑制剂,已经在临床验证中显示出很明确的疗效。
Isocitratedehydrogenase1(NADP+),异柠檬酸脱氢酶1。
位于第2号染色体上。
IDH1催化异柠檬酸的氧化脱羧2-酮戊二酸。
胶质母细胞瘤与IDH1有关系。
IDH1突变往往是在低级别胶质瘤发展的第一步,IDH1基因突变在这些脑肿瘤形成的关键事件。
IDH1突变与生存期延长相关。
胶质母细胞瘤与野生型IDH1基因有只有1年的中位总生存期,而IDH1突变的胶质母细胞瘤患者有2年以上的中位总生存期。
位于第9号染色体。
JAK2是一个非受体酪氨酸激酶,是催乳素受体的下游,被催乳素受体激活。
在白血病人中有发现JAK2和TEL的融合基因,也发现有JAK2和PCM的融合基因
Tyrosine-proteinkinase。
位于第19号染色体上。
JAK3主要表达于免疫细胞,并且响应白细胞介素受体的磷酸化信息号再向下传递激活信号。
JAK3突变会导致免疫能力低下,而免疫力低下是肿瘤不被免疫排除的重要原因。
IDH2
Isocitratedehydrogenase,异柠檬酸脱氢酶2。
位于第15号染色体。
是三羧酸循环中的一个重要的酶,把异柠檬酸脱氢,变成α-酮戊二酸。
IDH酶通常将异柠檬酸转化为α-酮戊二酸盐(αKG)。
然而,与癌症(cancer)相关的IDH突变产生了一种酶,能够将αKG转化为2-羟戊二酸(2HG)。
2HG能够竞争性地抑制αKG的靶点,后者具有DNA和组蛋白脱甲基酶活性。
因此,2HG产物导致了基因表达中的变化,这些变化之前被报告能够导致受损的分化。
IDH2突变导致了分化抑制,并在不同的肿瘤背景下改变了DNA甲基化和过度增殖。
当用一种小分子化合物AGI-6780治疗IDH2突变的小鼠白血病AML细胞时,AML细胞停止了增生和分化。
KDR
Kinaseinsertdomainreceptor,也叫VEGFR-2(vascularendothelialgrowthfactorreceptor2,血管内皮生长因子2),。
位于4号染色体
KDR则主要介导内皮细胞的增殖,导致血管通透性的增高,并阻止内皮细胞凋亡,维持内皮细胞的存活,与胚胎期内皮细胞的分化有关
与癌症的关系:
肿瘤的生长和扩散依赖于血管生成。
肿瘤血管生成对于肿瘤的发生、发展、转移及预后具有重要作用,肿瘤血管生成是一系列复杂的调控过程,其中VEGF及其受体发挥着重要作用
Regorafenib是KDR的特异性抑制剂,并已得到FDA的批准,用于结肠癌
Foretinib是KDR的抑制剂,同时也是c-Met的抑制剂
lenvatinib,motesanib,Pazopanib等也是KDR的抑制剂
KIT(CD117)
Mast/stemcellgrowthfactorreceptor,又名:
CD117。
CD117是在造血干细胞表面表达的一种细胞因子受体。
这个受与与干细胞因子结合后形成一个二聚体,这个二聚体激活其内在的酪氨酸激酶的活性,然后在细胞内激活下游的信号途径
精细胞肿瘤中经常会有CD117的第17个外显子的突变,而且这种肿瘤中还常常有CD117的过表达。
CD117与白血病、黑色素瘤、胃肠道间质肿瘤相关
Gleevec(格列卫,Imatinib),如果病人有CD117在17号外显子的突变,Gleevec就会有效。
dasatinib和nilotinib也有应用
KRAS
V-Ki-ras2Kirstenratsarcomaviraloncogenehomolog。
位于12号染色体
KRAS是一个分子开关,一旦它打开,它就会活化多种分裂、增殖因子,例如:
c-Raf和PI3-kinase。
KRAS会和GTP结合,把GTP的最末一个磷酸基切掉,让它变成GDP。
在把GTP变成GDP后,KRAS就关闭了。
正常情况下,KRAS活化后,会马上失活。
但是KRAS基因突变后,KRAS蛋白持续保持活化状态,导致细胞持续增生
与结直肠癌的关系密切。
20%的非小细胞肺癌中有KRAS突变
与药物的关系:
KRAS是EGFR信号通路的下游结点。
如果KRAS发生持续活化突变,则针对EGFR的抑制剂药物,如:
易瑞沙(Iressa,gefitinb)和特罗凯(erlotinib,Tarceva),往往无效。
所以,用针对EGFR的靶向药物治疗前,建议检测KRAS的突变状态
MET(HGFR)
MNNGHOSTransforminggene,又名HGFR、c-MET。
位于7号染色体
Hepatocytegrowthfactor(HGF)是MET的配体,它是可激活MET。
被激活的MET启动一系列的生物反应,并会引起入侵性生长。
MET参与的信号通路:
RAS
PI3K
STAT
beta-catenin
Notch
异常的MET活性会导致血管的生长,并以此支持肿瘤的营养。
激活关键的生长通路
促进血管生长
促进细胞的分散,这也导致肿瘤的转移
MET基因倍培是透明细胞瘤的一个潜在生物标记物
有MET基因倍增的结直肠癌肿瘤,对抑EGFR的治疗有抗性
一系列的MET抑制剂被研发出来:
K252a
SU11274
PHA-665752
ARQ197
Foretinib
SGX523
MP470
HGF的抑制剂
HGF是已知唯一的MET配体,所以能够阻断HGF作用的物质,也可能会是有效的抑制MET活性的物质
MET的假底物
CGEN241是MET酶的借底物,对动物模型中,对阻止肿瘤的生长和转移有很有效
主动免疫治疗
白介素2(IL-2)被FDA批准用于肾细胞癌、和转移的黑色素瘤,这两种瘤都常常有MET活性失控的情况。
MLH1
MutLhomolog1。
位于3号染色体上。
MLH1是人类基因中与大肠杆菌MutL同源的一个基因,MutL在大肠杆菌中负责对DNA错配的修复。
MLH1基因的突变与遗传性非息肉结直肠癌有密切关系。
MPL(CD110)
Myeloproliferativeleukemiavirusoncogene,又名ThrombopoietinReceptor(促血小板生成素受体)、CD110。
位于1号染色体上。
MPL是一个细胞因子受体,与配体结合后,能把下游的信号蛋白(STAT、MAPK等)的酪氨酸残基磷酸化。
MPL与骨髓增生白血病有关系。
NOTCH1
Notchhomolog1。
位于3号染色体。
Notch1基因在发育过程中控制细胞的分化命运。
Notch1与白血病相关。
NPM1
Nucleophosmin,又名:
nucleolarphosphoproteinB23、numatrin。
位于5号染色体上。
NPM1与核仁核糖体蛋白相关,能与单链或双链的核酸相结合。
在多种肿瘤中都发现NPM1的上调、突变、或者染色体易位。
当NPM1高表达时,会抑制P53/ARF通路
NRAS
NeuroblastomaRASviraloncogenehomolog。
位于1号染色体。
与KRAS基因相近,对细胞的生长起调控作用。
也是GTP酶,可以将GTP分解成GDP。
与肿瘤的相关笥:
与多种肿瘤相关。
PDGFRA
Alpha-typeplatelet-derivedgrowthfactorreceptor,是PDGFR的a亚基。
是血小板衍生生长因子家族的受体,也是一个细胞表面的酪氨酸激酶。
激活MAPK通路
激活PI3K通路
与胃肠道间质瘤有关系
Crenolanib(CP-868596)是一种有效的,选择性的PDGFRα/β抑制剂,Kd为2.1nM/3.2nM,也有效抑制FLT3,对D842V突变型而不是V561D突变型敏感,作用于PDGFR比作用于c-Kit,VEGFR-2,TIE-2,FGFR-2,EGFR,erbB2,和Src选择性高100倍以上。
PIK3CA
phosphatidylinositol-4,5-bisphosphate3-kinase,catalyticsubunitalpha,是PIK3的α亚基。
P110α。
位于第3号染色体上。
PIK3激活PI3K/AKT/mTOR通路中的蛋白激酶B的活性。
PIK3是一个重要的信号通路的关键酶。
在许多肿瘤中,都存在PIK3的突变。
这些突变大多数是导致激酶的活性更高。
Wortmannin是一种PI3K抑制剂
PTEN
Phosphataseandtensinhomolog。
位于10号染色体。
是一个磷酸酯酶,专门使PIP3脱掉磷酸基团。
这个活性的作用是阻断AKT信号通路。
是一个抑癌基因。
可以抑制细胞的分裂生长,并可以导致细胞的凋亡。
在人类肿瘤中,PTEN是最常见的被丢失的基因。
在前列腺肿瘤中,70%都有丢失至少一个拷贝。
在胶质母细胞瘤、子宫内膜癌、和前列腺癌中经常发现PTEN的失活突变。
PTEN的突变也会导致各种癌变倾向。
PTPN11
Tyrosine-proteinphosphatasenon-receptortype11,又名Shp2。
位于12号染色体。
PTPN11是一个酪氨酸磷酸酯酶。
它可以把结合在酪氨酸残基上的磷酸基团水解下来。
带有PTPN11突变的患者,很容易得少年单核细胞白血病。
在神经母细胞瘤,黑色素瘤,急性髓细胞性白血病,乳腺癌,肺癌,结肠直肠癌中有发现PTPN11的活化突变。
有报道:
PTPN11既有促癌的作用,也有抑癌的作用
RB1
retinoblastomaprotein。
位于第13号染色体
这是一种抑癌基因,可以抑制E2F因子的作用,而E2F-DF是一种把细胞推进到S期的重要因子。
这也是一个染色体重新组织的因子,它可以召集甲基化酶和乙酰化酶。
如果两个RB1等位基因都突变了,这人就会视网膜母细胞瘤,并因此瞎掉。
RET
RETproto-oncogene。
位于10号染色体上。
RET是一个受体酪氨酸激酶,参与神经的形成。
缺少这个基因的人会得先天性巨结肠症(因为肠子没有神经控制,就不会蠕动,大便把肠子撑得很大)。
与肿瘤的关系:
RET功能增强性的突变,会导致:
甲状腺髓样癌,多发性内分泌瘤形成(2A型,和2B型),嗜铬细胞瘤和甲状旁腺增生
凡德他尼(vandetanib)是EGFR、VEGFR和RET酪氨酸激酶的抑制剂。
卡博替尼(Cabozantinib)是VEGFR2,Met,FLT3,Tie2,KitandRet的强效抑制剂。
SMAD4
Mothersagainstdecapentaplegichomolog4。
位于第18号染色体上。
SMAD4和SMAD1、SMAD2等因子相结合,型成复合物,结合到DNA上以转录因子的形式起作用。
在结直肠癌和胰腺癌中经常发现有SMAD4的突变。
它也在常染色体显性遗传病幼年性息肉综合症中被发现有突变,这些息肉很可以发展成结肠癌。
SMARCB1
SWI/SNF-relatedmatrix-associatedactin-dependentregulatorofchromatinsubfamilyBmember1。
位于第22号染色体。
这个基因所编码的蛋白,是解开染色体压缩结构的复合蛋白的一个组成部分。
它能使转录复合体能够更高效地结合到目标基因区域上。
将编码核蛋白质也可结合并增强HIV-1整合酶对DNA的结合活性。
这是一个肿瘤抑制基因,与恶性横纹肌样瘤有关系。
SMO
S
 升级会员
升级会员 