第九章细胞骨架知识点Word文件下载.docx
《第九章细胞骨架知识点Word文件下载.docx》由会员分享,可在线阅读,更多相关《第九章细胞骨架知识点Word文件下载.docx(13页珍藏版)》请在冰豆网上搜索。
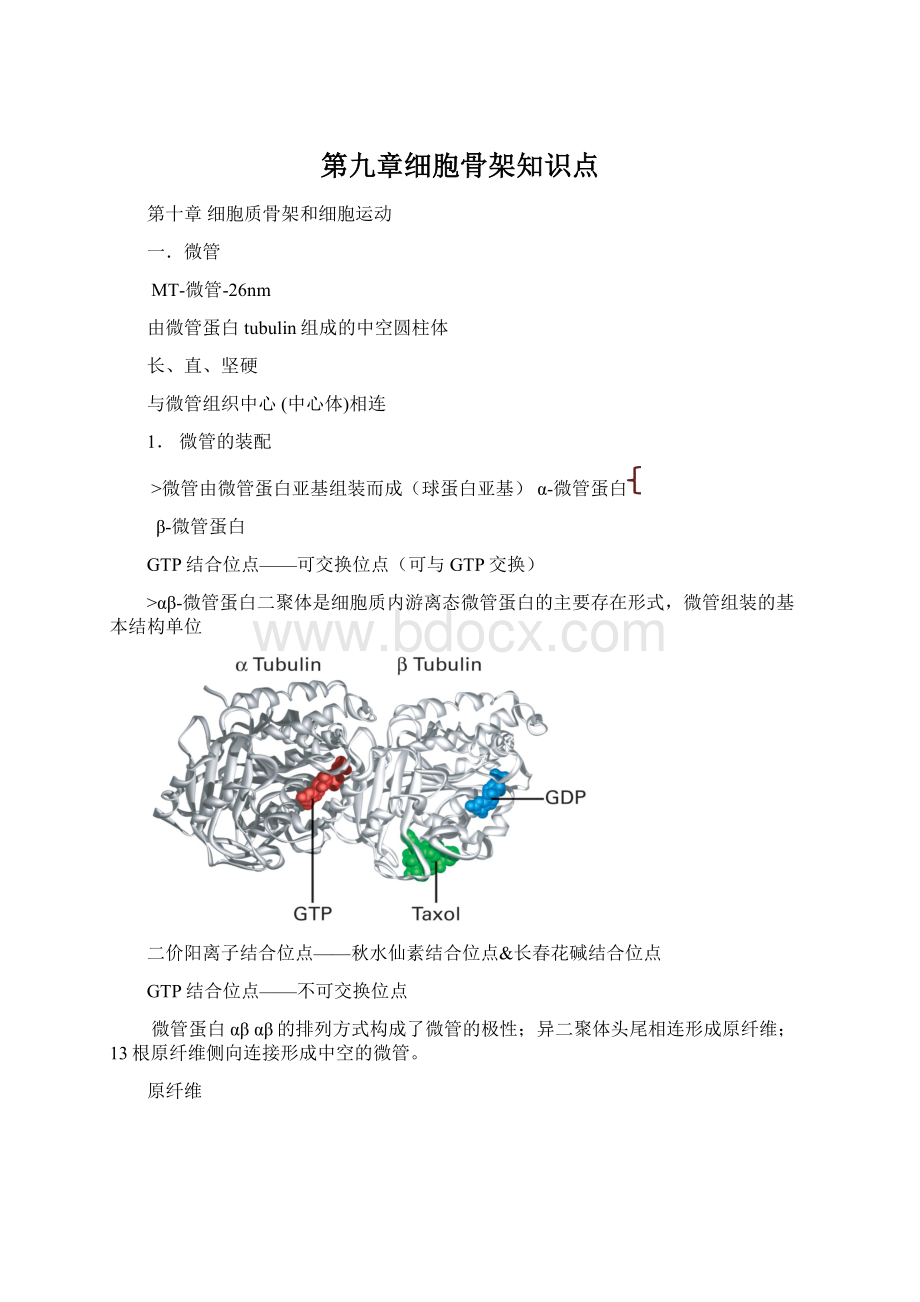
13根原纤维侧向连接形成中空的微管。
原纤维
踏车行为
微管(+)极的装配速度快于(—)极的装配速度;
或微管一端发生装配使微管延长,而另一端发生去装配使微管缩短,这种现象称为踏车行为。
微管装配的条件:
微管蛋白浓度、GTPcap和温度
当二聚体浓度低于临界浓度(Cc),则微管解聚
当二聚体浓度高于临界浓度,则组装微管
因为Cc(负极)>
Cc(正极),所以正极装配快于负极
当Cc(正极)<
C<
Cc(负极)时,则正极装配,负极解聚,
即踏车现象。
微管体外装配影响因素
聚合:
微管蛋白浓度≥1mg/mL
(二聚体蛋白浓度大于纤维状蛋白浓度)、
370C、有Mg2+、有GTP供应、低Ca2+
解聚:
低温、高压、高Ca2+
2.微管组织中心(MTOC-microtubuleorganizingcenter)
微管在生理状态以及实验处理解聚后重新装配的发生处称为微管组织中心。
多数微管的一端固着MTOC,如基体或中心体。
基体
MTOC决定微管的极性,负极指向MTOC,正极背向MTOC。
微管
中心粒(桶状结构)
每个中心体含有一对中心粒(彼此垂直分布)
单管、双联管(鞭毛、纤毛)和三联管(中心粒、基体)
3.微管结合蛋白(MAPs和τ蛋白)
作用
(1)稳定微管的空间结构
(2)促使微管蛋白/微管的动态平衡趋于装配
Tau蛋白
4.微管特异性药物
秋水仙素、长春花碱:
阻止装配
紫杉醇:
阻止解聚
5.功能
(1)维持细胞形态
(2)细胞内物质运输--------颗粒和囊泡
细胞内的物质运输需要马达蛋白的带动;
马达蛋白:
驱动蛋白或动力蛋白(正极或负极指向,ATP)
与微管结合的马达蛋白:
利用ATP水解酶释放的能量驱动自身沿微管定向运动的蛋白。
与ATP结合时,两个头部均结合在微管上;
与ADP结合时,则一个头部结合在微管上。
ATP的水解引起:
可逆的构象改变;
单方向的移动
驱动蛋白(Kinesin):
正极方向
它们的头部是相似的马达结构域,与ATP结合,在微管上移动;
它们的尾部结构域是不同的,用于带动不同的物质移动。
动力蛋白(dynein):
负极方向
动力蛋白由多种蛋白组成
沿微管向微管的负端移动,(胞质)动力蛋白可以将细胞器从细胞边缘向细胞中央转移。
如:
胞内体、溶酶体、ER膜泡等
驱动蛋白受体
囊泡
驱动蛋白
(3)鞭毛和纤毛运动
9+2结构,鞭毛滑动机制(动力蛋白臂)
纤毛和鞭毛是由质膜包围,且突出于细胞表面、由微管和动力蛋白等构成的高度特化的细胞结构。
(4)有丝分裂中纺锤体的形成和染色体移动
(5)形成基体和中心体
二.微丝(microfilament)
又称肌动蛋白丝/纤维状肌动蛋白,是由肌动蛋白(actin)组成的直径为7nm的骨架纤维。
主要结构成分是肌动蛋白
肌动蛋白内部有一个核苷酸(ATP或ADP)和一个二价阳离子(Mg2+或Ca2+)的结合位点
6种肌动蛋白α-肌动蛋白横纹肌
心肌
血管平滑肌
肠道平滑肌
β-肌动蛋白
γ-肌动蛋白
1微丝的装配
以3个肌动蛋白单体为核心
(+)极装配比(–)极快
肌动蛋白单体具有极性,装配时呈头尾相接,故微丝也具有极性,具有踏车现象。
微丝装配的条件:
肌动蛋白浓度、ATPcap
因为Cc(负极)>
Cc(负极)时,则正极装配,负极解聚,即踏车现象。
在体内,有些微丝是永久性的结构,通常微丝是一种动态结构,不断进行装配和解聚。
微丝的装配条件
体外装配
解聚:
有Ca2+和低浓度的Na+、K+阳离子
组装:
有Mg2+和高浓度的Na+、K+阳离子
体内装配
在体内,有些微丝是永久性的结构,如肌细胞中的细丝和小肠上皮细胞微绒毛中的微丝等。
有些微丝是暂时性的结构,如胞质分裂时的收缩环、血小板激活时丝状突起中的微丝等。
通常微丝是一种动态结构,不断进行装配和解聚。
2微丝结合蛋白(MFAP)
肌动蛋白单体结合蛋白:
阻止单体肌动蛋白组装成纤维状肌动蛋白
(胸腺素β4、前纤维蛋白)
MF的不同存在形式与MFAP的种类有关,MFAP参与MF高级结构的形成,并调节MF的装配。
如参与微丝与膜的结合;
可横向连接微丝形成束;
结合于纤维一端,阻止肌动蛋白单体的增加或减少;
将微丝切断;
成核作用(ARP);
将微丝交联成网状分布等
微丝结合蛋白的作用
成核,促使微丝装配(ARP2/3)
与肌动蛋白单体结合,阻止或促使微丝装配(胸腺素thymosin,前纤维蛋白profilin)
与微丝末端结合(CapZ)
交联作用,使微丝呈束状或网状排列(丝束蛋白、α-辅肌动蛋白、细丝蛋白)
截断和封端作用(凝溶胶蛋白gelsolin,切丝蛋白cofilin)
使微丝与细胞膜相连接(形成蛋白formins)
3.微丝特异性药物
细胞松弛素:
切断微丝,并结合在微丝末端阻止聚合
鬼笔环肽:
稳定微丝,抑制解聚
4.功能
(1)肌肉收缩
粗丝(肌球蛋白)和细丝(肌动蛋白)的相对滑动机制
肌球蛋白是微丝结合蛋白,也是沿微丝运动的马达蛋白(+极方向)
粗肌丝由肌球蛋白组装而成,肌球蛋白的头部突出于粗肌丝的表面,具有ATP酶活性,构成粗丝的横桥,与肌动蛋白分子结合。
细肌丝的主要成分是肌动蛋白,辅以原肌球蛋白和肌钙蛋白。
肌肉收缩的机制
①产生动作电位,使肌细胞质膜去极化,释放Ca2+至肌浆中,钙离子浓度升高;
②钙离子与Tn-C结合,引起肌钙蛋白构象变化,使肌动蛋白与Tn-T脱离,并解除肌动蛋白与肌球蛋白结合的障碍;
③当肌球蛋白的头部结构域没有与ATP结合,该肌球蛋白的头部与细肌丝结合,处于僵直状态;
④当ATP结合到肌球蛋白的头部,引起头部结构域与细肌丝的分离;
⑤ATP被头部结构域的ATP酶水解成ADP与Pi,使肌球蛋白构象变化发生旋转,向细肌丝的正极端抬升,并结合到靠近细肌丝正极端的一个肌动蛋白亚基上;
⑥Pi释放,肌球蛋白颈部结构域发生构象变化,拉动细肌丝导致细肌丝与粗肌丝的滑动;
⑦ADP释放,肌球蛋白的头部结构域与细肌丝之间又回到僵直状态;
如果体系中仍有高浓度的钙离子,则继续下一周期;
若到达肌细胞的冲动一旦停止,肌质网就通过钙泵回收钙离子,收缩周期停止。
(2)变形运动、吞噬运动
收缩环
微绒毛平行的微丝,微丝无收缩功能,起支撑作用
(3)胞质环流在植物细胞和其他细胞中,细胞质的流动是围绕中央液泡进行的环形流动模式
(4)粘着带和粘着斑
三.中间纤维(intermediatefilament)
1.中间纤维的装配
(1)二聚体:
在α螺旋区形成双股超螺旋。
(2)四聚体:
2个二聚体反向平行、1/4分子交叠排列,四聚体是IF解聚的最小单位。
(3)8个四聚体装配成IF。
IF没有极性,没有特异性药物;
游离单体很少。
也没有典型的踏车行为
①由2个中间纤维蛋白亚基平行排列组装而成双股螺旋的二聚体;
②两个二聚体按反向平行、半分子交叠的方式组装成四聚体;
③四聚体首尾相连形成原纤维;
④8根源纤维构成圆柱状的10nm纤维;
2.Lamina与IF
有许多共同点:
均形成10nm纤维;
都能抵抗高盐和非离子去污剂的抽提;
抗体反应相同;
蛋白结构相似。
3.中间纤维结合蛋白(IFAP)
约有15种IFAP,功能多样,如使中间纤维形成聚集物;
在IF、MT和MF之间形成横桥;
参与IF与膜的结合;
桥板蛋白使IF与桥粒连接
4.功能
IF在细胞质中起支架作用,并与细胞核定位有关,也在细胞间或组织中起支架作用;
在细胞质中与MT、MF和细胞器构成一个完整的网架系统;
在细胞质外侧与膜和胞外基质直接联系;
在细胞质内部与核膜和核基质直接联系。
核功能-------信息分子或信息分子前体
IF在体外与单链DNA高度亲和;
与H1以外的其他组蛋白高度亲和;
组蛋白不能被Ca2+依赖的蛋白酶水解,但与IF结合后可被水解,可导致DNA双螺旋发生解旋。
 升级会员
升级会员 