高中生物竞赛辅导资料细胞生物学.docx
《高中生物竞赛辅导资料细胞生物学.docx》由会员分享,可在线阅读,更多相关《高中生物竞赛辅导资料细胞生物学.docx(44页珍藏版)》请在冰豆网上搜索。
高中生物竞赛辅导资料细胞生物学
高中生物竞赛辅导资料:
第一章细胞生物学
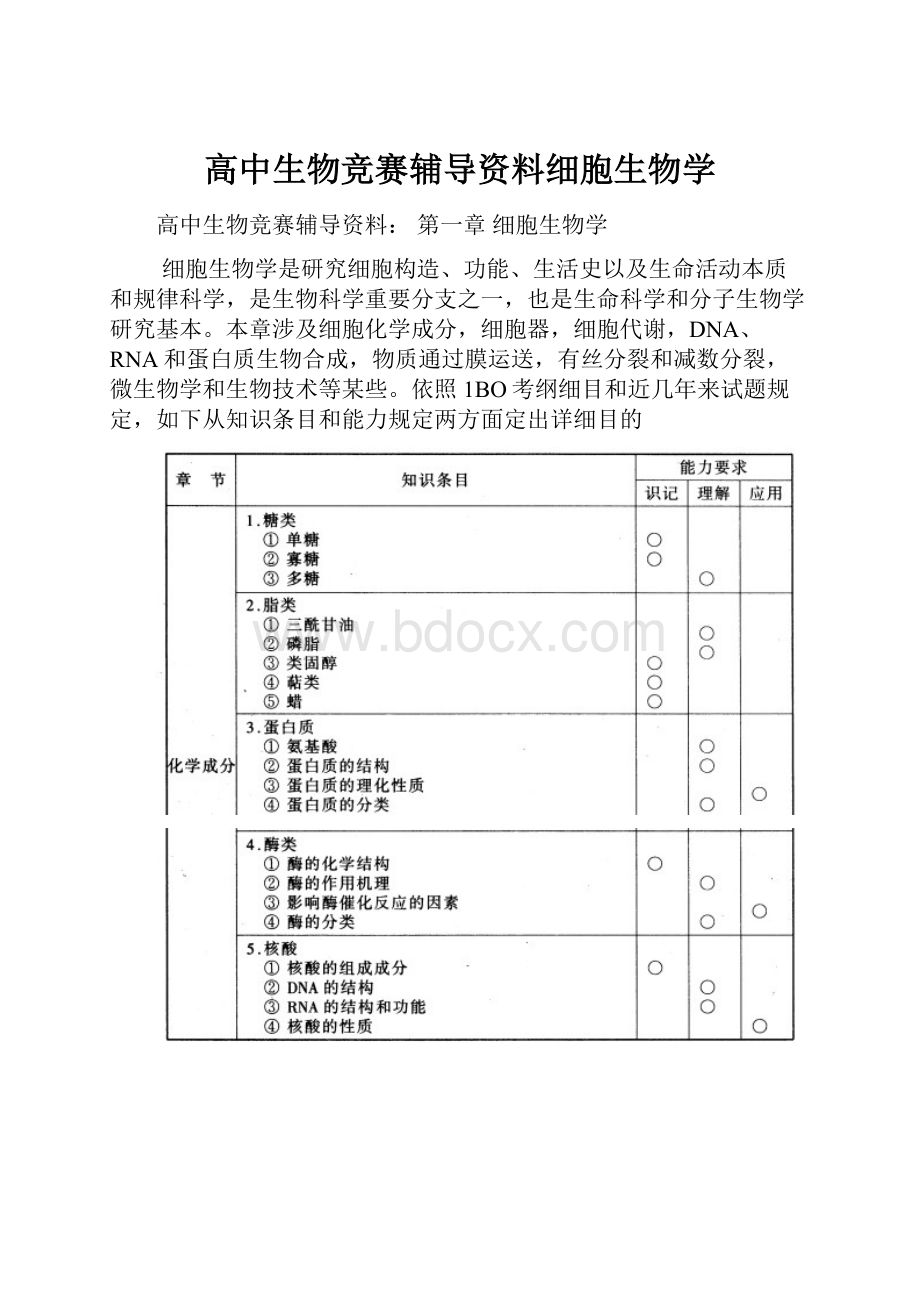
细胞生物学是研究细胞构造、功能、生活史以及生命活动本质和规律科学,是生物科学重要分支之一,也是生命科学和分子生物学研究基本。
本章涉及细胞化学成分,细胞器,细胞代谢,DNA、RNA和蛋白质生物合成,物质通过膜运送,有丝分裂和减数分裂,微生物学和生物技术等某些。
依照1BO考纲细目和近几年来试题规定,如下从知识条目和能力规定两方面定出详细目的
第一节细胞化学成分
尽管自然界细胞形态多样,功能各异,但其化学成分基本相似,重要涉及:
糖类、脂类、蛋白质、核酸、酶类等。
一、糖类
糖类是多羟基醛、多羟基酮总称,普通可用Cm(H20)n化学通式表达。
由于某些糖分子中氢和氧原子数之比往往是2:
1,与水构造相似,故又把糖类称为碳水化合物。
糖是生命活动重要能源,又是重要中间代谢物,尚有些糖是构成生物大分子,如核酸和糖蛋白成分,因而具备重要意义。
糖类化合物按其构成可分为单糖、寡糖、多糖。
如果糖类化合物中尚具有非糖物质某些,则称为糖复合物,例如糖蛋白、蛋白多糖、糖脂和脂多糖等。
(一)单糖
单糖是最简朴糖,不能被水解为更小单位。
单糖普通具有3—7个碳原子,分别称为丙糖、丁糖、戊糖、己糖和庚糖。
天然存在单糖普通都是D-构型。
单糖分子既可以开链形式存在,也可以环式构造形式存在。
在环式构造中如果第一位碳原子上羟基与第二位碳原子羟基在环伺一面,称为α-型;如果羟基是在环两面,称β-型。
重要单糖有如下几种:
1.丙糖如甘油醛(醛糖)和二羟丙酮(酮糖)。
它们磷酸酯是细胞呼吸和光合伙用中重要中间代谢物。
2.戊糖戊糖中最重要有核糖(醛糖)、脱氧核糖(醛糖)和核酮糖(酮糖)。
核糖和脱氧核糖是核酸重要成分,核酮糖是重要中间代谢物。
3.己糖葡萄糖、果糖和半乳糖等都是己糖。
所有己糖分子式为C6H1206,但构造式不同,互为同分异构体。
葡萄糖是植物光合伙用产物,也是细胞重要能源物质之一。
(二)寡糖
由少数几种(2—6个)单糖缩合而成糖称为寡糖。
最多寡糖是双糖,如麦芽糖、蔗糖、纤维二糖、乳糖。
1.麦芽糖麦芽糖是由一种α—D-葡萄糖半缩醛羟基与另一分子α-D-葡萄糖C4上醇羟基缩合脱去一分子水,通过α-1,4-糖苷键结合而成。
麦芽糖是淀粉基本单位,淀粉水解即产生麦芽糖,因此麦芽糖普通只存在于淀粉水解组织,如麦芽中。
2.蔗糖一分子α-D—葡萄糖和一分子β-D-果糖缩合脱水即成蔗糖。
甘蔗、甜菜、胡萝卜以及香蕉、菠萝等水果中都富含蔗糖。
3.乳糖乳糖由一分子β-D-半乳糖和一分子α-D-葡萄糖通过β-1,4-糖苷键结合而成。
乳糖重要存在于哺乳动物乳汁中。
4.纤维二糖纤维二糖是纤维素基本构造单位,由2分子p-D-葡萄糖通过β-1,4-糖苷键结合而成。
(三)多糖
自然界数量最大糖类是多糖。
多糖是由诸多单糖分子缩合脱水而成分支或不分支长链分子。
常用多糖有:
淀粉、纤维素、糖原、几丁质和黏多糖等。
1.淀粉天然淀粉由直链淀粉与支链淀粉构成。
直链淀粉是α-D-葡萄糖基以α-1,4-糖苷键连接多糖链。
支链淀粉分子中除有α—1,4-糖苷键糖链外,尚有α-1,6-糖苷键连接分支。
淀粉与碘有呈色反映,直链淀粉为蓝色,支链淀粉为紫红色。
在稀酸或酶作用下,淀粉水解:
淀粉→糊精→麦芽糖→α-D—葡萄糖。
糊精是淀粉水解最初产物,随着水解,糖分子逐渐变小,它与碘作用分别呈红色、黄色、无色。
这个反映可用于淀粉水解过程检查。
2.糖原糖原是动物组织中贮存多糖,又称动物淀粉。
糖原也是α-D-葡萄糖基以α-1,4-糖苷键连接而成,但糖原分支比支链淀粉多。
糖原遇碘作用呈红褐色。
3.纤维素,纤维素是一种线性由β-D-葡萄糖基以β-1,4-糖苷键连接没有分支同多糖。
纤维素是植物细胞壁重要构成成分。
4.几丁质(甲壳素)昆虫和甲壳类外骨骼重要成分为几丁质,是N-乙酰—D-氨基葡萄糖以β-1,4-糖苷键缩合成同多糖。
二、脂类
脂类是生物体内一类重要有机化合物。
它们有一种共同物理性质,就是不溶于水,但能溶于非极性有机溶剂(如氯仿、乙醚、丙酮等)。
脂类构成元素重要有C、H、0,但0元素含量低,C、H元素含量高,彻底氧化后可以放出更多能量。
此外,有脂类还具有P和N。
生物体内常用具备重要生理功能脂类重要有三酰甘油、磷脂、类固醇、萜类、蜡等。
1.三酰甘油
三酰甘油也称脂肪,是由1分子甘油和3分子脂肪酸结合而成酯。
右边构造式中Rl、R2、凡是脂肪酸烃基链,构成三酰甘油脂肪酸可分为饱和脂肪酸和不饱和脂肪酸。
饱和脂肪酸碳氢链上没有双键,如软脂酸、硬脂酸,其熔点高。
不饱和脂肪酸碳氢链上具有不饱和双键,如油酸含1个双键,亚油酸含2个双键,亚麻酸含3个双键,因而熔点较低。
动物脂肪大多富含饱和脂肪酸,在室温下为固态,植物油含大量油酸和亚油酸,在室温下为液态。
对于哺乳动物和人,亚油酸和亚麻酸不能自己合成,只能从外界摄取,称为必须脂肪酸。
2.磷脂
磷脂又称甘油磷脂,此类化合物是甘油第三个羟基被磷酸所酯化,而其她两个羟基被脂肪酸酯化。
磷脂酸是最简朴磷脂,是其她复杂磷脂中间产物。
若磷脂酸分子中H为胆碱、胆胺、丝氨酸所取代,则分别成为卵磷脂、脑磷脂、丝氨酸磷脂等。
磷脂分子由于有磷酸及与之相连含氮化合物,因而是有极性分子:
它有磷酸一端为极性头,是亲水,它2个脂肪酸链为非极性尾,是疏水。
如将磷脂放在水面上,磷脂分子都将以亲水头和水面相接,而倒立在水面上,成一单分子层。
如将磷脂放入水中,磷脂分子则会形成单分子微团,各分子极性头位于微团表面而与水接触,非极性疏水端则藏在微团中心。
3.类固醇
类固醇分子基本构造是环戊烷多氢菲。
最熟知类固醇是在环戊烷多氢菲上连有一种碳氢链胆固醇。
胆固醇是动物膜和神经髓鞘重要成分,与膜透性关于。
性激素、维生素D和肾上腺皮质激素都属于类固醇。
4.萜类
萜类是由不同数目异戊二烯连接而成分子。
维生素A(视黄醇)、维生素E、维生素K、类胡萝卜素都是萜类。
β-类胡萝卜素裂解就成2个维生素A,维生素A可氧化成视黄醛,对动物感光活动有重要作用。
5.蜡
蜡是由高碳脂肪酸和高碳醇或固醇所形成脂,它存在于皮肤、毛皮、羽毛、树叶、昆虫外骨骼中,起保护作用。
三、蛋白质
蛋白质是细胞和生物体重要构成成分,普通占细胞干重一半以上。
蛋白质重要由C、H、0、N四种元素构成,其中氮含量在各种蛋白质中比较接近,平均为16%,因而用凯氏(KJelahl)法定氮测定蛋白质含量时,受检物质中含蛋白质量为氮含量6.25倍。
蛋白质是高分子化合物,其基本构成单位是氨基酸。
(一)氨基酸
1.氨基酸构造
天然存在于蛋白质中氨基酸共有20种,各种氨基酸(除脯氨酸)在构造上一种共同特点是,在与羧基相连碳原子(α-碳原子)上均有一种氨基,因而称为α-氨基酸,它们不同之处在于侧链,即R基不同。
除甘氨酸外,所有氨基酸分子中α-碳原子都是不对称,有L-型和D-型之分。
在天然蛋白质中存在氨基酸都是L-α-氨基酸。
2.氨基酸分类
依照R基团极性不同,氨基酸可分为:
非极性氨基酸(9种);极性不带电荷氨基酸(6种);极性带负电荷氨基酸(2种);极性带正电荷氨基酸(3种)。
如表1-1-1所示。
依照成年人营养需求,20种氨基酸又可分为必须氨基酸和非必须氨基酸。
必须氨基酸足指成年人体内不能合成而必要山食物提供一类氨基酸,涉及亮氨酸、异亮氨酸、缬氨酸、另;氨酸、蛋氨酸、色氨酸、赖氨酸、苯丙氨酸等8种。
精氨酸和组氨酸,在幼儿时期体内合成量满足不了生长需要,需食物补充,称为半必须氨基酸。
3.氨基酸重要理化性质
(1)普通物理性质α-氨基酸呈无色结晶,在水中溶解度各不相似,易溶于酸、碱,但不溶于有机溶剂、
(2)两性解离和等电点α-氨基酸在中性水溶液中或固体状态下重要是以两性离子形式存在,即在同一种氨基酸分子上带有能放出质子-NH3+正离子和能接受质子
@一C00-负离子。
因而,氨基酸是两性电解质。
当两性离子氨基酸溶解于水时,其正负离子都能解离,但解离度与溶液pH值关于。
向氨基酸溶液加酸时,其两性离子-COO-负离子接受质子,自身成为正离子,在电场中向阴极移动加入碱时,其两性离子一NH3+正离子解离放出质子(与一OH-合成水),其自身成为负离子,在电场中向阳极移动。
当凋节氨基酸溶液pH值,使氨基酸分子上一NH3+和一C00-解离度完全相等时,即氨基酸所带净电荷为零,在电场中既不向阳极移动也不向阴极移动,此时氨基酸所处溶液pH值称为该氨基酸等电点,以符号pI表达。
在等电点时,氨基酸溶解度最小,容易沉淀,运用这一性质可以分离制备各种氨基酸。
(3)紫外吸取光谱各种氨基酸在可见光区都没有光吸取,在远紫外区均有光吸取,而在近紫外光区仅色氨酸、酪氨酸、苯丙氨酸有吸取能力。
其中色氨酸最大吸取波长为279nm,酪氨酸最大吸取波长278nm,苯丙氨酸最大吸取波长为259nm。
运用紫外光法可以测定这些氨基酸含量。
(4)重要化学反映氨基酸不但α-氨基、α-羧基能参加反映,并且有侧链R基团也能参加化学反映,因而可以发生反映诸多。
如:
α-氨基能与茚三酮反映产生蓝紫色沉淀(脯氨酸和羟脯氨酸则产生黄色沉淀);α-氨基可与亚硝酸反映产生氮气,在原则条件下测定氮气体积,即可计算出氨基酸量;某些氨基酸R基团能与特殊试剂发生呈色反映。
(二)蛋白质构造
已确认蛋白质构造有不同层次,人们为了结识以便普通将其分为一级构造、二级构造,超二级构造、构造域、三级构造及四级构造。
l,一级构造
蛋白质一级构造又称为初级构造或化学构造,是指蛋白质分子内氨基酸排列顺序。
蛋白质分子中氨基酸重要通过肽键互相连接。
肽键是由一种氨基酸分子中α-氨基与相邻另一种氨基酸分子中α-羧基,通过缩水而成,这样连起来氨基酸聚合物叫做肽。
多肽链上各个氨基酸由于在互相连接过程中丢失了α-氨基上H和α-羧基上OH,被称为氨基酸残基。
在多肽链一端氨基酸具有一种未反映游离氨基(一NH2),称为肽链氨基末端氨基酸或N末端氨基酸,另一端氨基酸具有一种尚未反映游离羧基(一COOH),称为肽链羧基末端氨基酸或C末端氨基酸。
普通表达多肽时,总是N末端:
写在左边,C末端写在右边。
肽链中除肽键外尚有二硫键,它是由肽链中相应部位上两个半胱氨酸脱氢连接而成,是肽链内和肽链间重要桥键。
2,二级构造
二级构造是指多肽链自身绕曲折叠成有规律构造或构象。
这种构造是以肽链内或肽链间氢键来维持。
常用二级构造有α-螺旋、β-折叠、β-转角、自由绕曲等四种。
(1)α-螺旋α-螺旋模型是Pauling和Corey等研究羊毛、马鬃,猪毛、鸟毛等α-角蛋白时提出。
如图l-1-1所示,其特性是:
多肽链中氨基酸残基以100°角度环绕螺旋轴心盘旋上升,每3.6个残基就旋转一圈,螺距为0.54nm,即每个残、基沿螺旋体中心轴上、升0.15nm;右手旋转;多肽链内氢键由肽链中一种肽键一CO氧原子与第四个肽键一NH氢原子构成,每个氢键所形成环内共有13个原子,这种螺旋称为3.613.一条多肽链能否形成α-螺旋以及形成螺旋体稳定限度与R基团大小、带电状况等关于。
如多聚赖氨酸在pH7.0时,R基团带正电互相排斥,破坏螺旋形成,而在pH12时则能自发形成α-螺旋。
又如肽链内相邻残基是异亮氨酸、缬氨酸、亮氨酸等时,由于R基团较大,会阻碍α-螺旋形成。
多聚脯氨酸则由于肽键上不具备亚氨基氢,无法形成氢键,因而多肽链中只要有脯氨酸残基,α-螺旋即被中断,使多肽主链产生一种“结节”。
(2)β-折叠分两种类型,一是平行式,即所有肽链N端都在同一端,另一类是反平行式,即肽链N端一顺一反地排列。
β-折叠构造肽链几乎是完全伸展,邻近两链以相反或相似方向平行排列成片层状。
两个氨基酸残基之间轴心距离为0.35nm,β-折叠构造氢键是由两条肽链中一条一C0基与另一条一NH基形成。
丝蛋白二级构造重要是β-折叠。
如图1-1-2所示。
(3)β-转角蛋白质分子多肽链上经常浮现180°回折,在这种肽链回折角上就是β-转角构造,由第一种氨基酸残基一CO与第四个氨基酸残基一NH形成氢键。
(4)自由绕曲是指没有一定规律松散构造,酶功能部位经常处在这种构象区域中。
3.超二级构造与构造域
近年在研究蛋白质构象、功能与进化时,引进了超二级构造和构造域(图1-1-3)构造层次。
它们是二级构造与三级构造过渡型构象。
超二级构造是指若干相邻二级构造中构象单元彼此互相作用,形成有规则、在空间上能辨认二级构造组合体。
普通有βαβ、βββ、αα、ββ等。
如肌球蛋白、原肌球蛋白和纤维蛋白原中有一种αα超二级构造,是由两股或三股右手α-螺旋彼此缠绕而成左手螺旋构象。
构造域是指多肽链在超二级构造基本上进一步绕曲折叠成紧密球状构造,在空伺上彼此分隔各自具备某些生物功能亚构造。
普通状况下,酶活性部位位于两个构造域之间裂缝中。
4.三级构造
纤维状蛋白质普通只有二级构造,而球状,蛋白质在二级构造基本上,通过超二级构造和构造域,进一步组装成三级构造(图1-1-4)。
维持三级构造作用力重要是某些次级键,涉及氢键、盐键、疏水键和范德华力等。
其中疏水键在维持蛋白质三级构造上有突出作用。
5.四级构造
四级构造(图1-l-5)是指蛋白质分子内具备三级构造亚单位通过氢键、盐键、疏水键和范德华力等弱作用力聚合而成特定构象。
所谓亚单位,又称亚基,是指那些在化学上相互独立但自身又具备特定构象共同构成同一蛋白质肽链。
如血红蛋白有四个不同亚基,这4个亚基以一定形式结合在一起,形成特定构象,即是四级构造。
(三)蛋白质理化性质
1.胶体性质蛋白质相对分子质量很大,在水溶液中所形成颗粒具备胶体溶液特征,如布朗运动、丁达尔现象、不能通过半透膜等。
溶液中,蛋白质胶体颗粒带有相似电荷,彼此排斥;并且颗粒表面极性分子能与水分子形成一层水膜,将蛋白质颗粒互相隔开,因而蛋白质颗粒比较稳定,不易沉淀。
2,两性电解质蛋白质分子除了肽链两端有自由α-氨基和α-羧基外,许多氨基酸残基侧链上存有不少可解离基团,因此蛋白质是两性电解质。
在酸性溶液中蛋白质带正电,在碱性淀液中蛋白质带负电。
当溶液达到某一pH值时,蛋白质所带正负电荷相等,这时溶液pH值叫做蛋白质等电点(pI)。
普通含酸性氨基酸较多蛋白质,等电点偏酸;含碱性氨基酸较多蛋白质,等电点偏碱。
可以依照不同蛋白质等电点,用电泳法分离蛋白质。
3.沉淀反映如果在蛋白质溶液小加入恰当试剂,破坏了蛋白质水膜或中和蛋白质电荷,则蛋白质胶体溶液就不稳定而浮现沉淀现象.可引起沉淀反映试剂有高浓度盐类(如硫酸铵、硫酸钠、氯化钠等,称为盐析),有机溶剂(如酒精、丙酮),重金属盐(如硝酸银、醋酸铅、三氯化铁等),某些生物碱试剂(如苦味酸、单宁酸等).
4.变性蛋白质因受物理或化学因素影响,其分子空间构造变化,导致其理化性质、生物活性都发生变化,这种现象称为蛋白质变性。
能使蛋白质变性化学因素有强酸、强碱、重金属离子、尿素、酒精、丙酮等;能使蛋白质变性物理因素有加热震荡或搅拌、超声波、紫外线及X射线照射等。
蛋白质生物活性丧失是蛋白质变性重要特性,变性后蛋白质最明显理化性质变化是溶解度减少。
变性过程中不发生肽键断裂和二硫键破坏,因而不发生一级构造破坏;而重要发生氢键、疏水键破坏,使肽链有序卷曲、折叠状态变为松散无序。
5.紫外吸取蛋白质在280nm紫外光下,有最大吸取峰。
这重要是由于肽链中酪氨酸和色氨酸R基团引起。
因而可以用紫外线分光光度法测定蛋白质在280nm光吸取值来测定蛋白质含量。
6.变构作用含2个以上亚基蛋白质分子,如果其中一种亚基与小分子物质结合,那么不但该亚基构象发生变化,并且其她亚基构象受影响也发生变化,成果整个蛋白质分子构象乃至活性均会变化,这一现象称为变构作用(或别构作用)。
例如,血红蛋白有4个亚基,当02与其中一种亚基结合后,即引起该亚基构象变化,进而又会引起此外三个亚基构象发生变化,成果整个分子构象变化,使所有亚基更易于与氧结合,大大加快血红蛋白与氧结合速度。
7.呈色反映蛋白质分子中因具有某些特殊构造或某些特殊氨基酸残基,能与各种化合物发生颜色反映。
重要颜色反映如表l-1-2所示。
(四)蛋白质分类
1.蛋白质化学分类依照蛋白质分子构成可将蛋白质分为简朴蛋白质和结合蛋白质两大类。
简朴蛋白质完全水解产物为α-氨基酸,即只由α-氨基酸构成。
因而,简朴蛋白,质又称单纯蛋白质,如球蛋白、白蛋白、组蛋白等。
结合蛋白质由简朴蛋白质和非蛋白质物质两某些构成。
非蛋白质某些普通称为辅基。
辅基可以是核酸、糖类、脂类、色素、磷酸,由此构成结合蛋白质分别称为核蛋白、糖蛋白、脂蛋白、色蛋白、磷蛋白。
2.蛋白质功能分类依照蛋白质功能大体分为构造蛋白和酶两大类,构造蛋白参加细胞构造构成。
酶是活细胞产生具极高催化效率一类蛋白质,生物体内绝大多数化学反映都需要在酶催化作用下才干进行。
四、酶类
酶是由活细胞产生,能在体内或体外起同样催化作用一类具备活性中心和特殊构象生物大分子,涉及蛋白质和核酸。
生物体和细胞内错综复杂彳弋谢反映必要具备酶才干按一定规律有条不紊地进行。
酶缺陷或者酶活性被抑制会引起生物体和细胞病变。
在这里重要讨论蛋白质属性酶。
(一)酶化学构造
绝大多数酶是蛋白质,依照酶化学构成.可以把酶提成单纯酶和结合酶,单纯酶分子完全由蛋白质构成,不含其她成分。
结合酶分子由简朴蛋白质(称为酶蛋白)和辅助因子两某些构成,辅助因子可以是金属离子或小分子有机物。
普通把这些小分子有机物称为辅酶或辅基。
辅酶指与酶蛋白结合比较松,用透析法可以除去小分子有机物;而辅基则指与酶蛋白结合比较紧,用透析法不易除去小分子有机物;两者没有本质区别。
酶催化反映专一性和高效性重要决定于酶蛋白。
酶分子中有诸多化学基团,但并不是所有基团都与酶活性关于。
酶活性仅与一某些基团有直接关系,这些基团称为酶必须基团。
如果对这些基团进行取代或修饰,则酶活性丧失。
酶必须基团在一级构造上也许相距很远,甚至也许不在一条肽链上,但由于肽链盘绕折叠使它们在空间上彼此接近,形成具备一定空间构造区域,而直接与酶催化功能关于,这种区域称为酶活性中心。
酶活性中心涉及两个功能部位:
一种是结合部位,一定底物靠此部位结合到酶分子上;一种是催化部位,底物分子中化学键在此处被打断或形成新化学键,从而发生一定化学反映。
(二)酶作用机制
1.酶催化作用——减少活化能
在一种反映体系中,任何反映物分子均有进行化学反映也许,但并非所有反映物分子都进行反映。
由于在反映体系中各反映物分子所含能量高低不同,只有那些所含能量达到或超过一定限度(称为能阈)活化分子(处在过渡态分子)才干在碰撞中发生化学反映。
显然,活化分子越多,反映速度越快。
将分子由常态转变到活化状态(过渡态)所需能量,称为活化能。
酶催化作用就是减少化学反映活化能,由于在酶催化反映中只需较低能量就可使反映物进入“过渡态”,因此同非酶催化反映相比,活化分子数量大大增长,从而加快反映速度。
如图1-1-6所示。
2.中间产物学说
酶为什么能减少反映活化能?
中间产物学说能比较好地解释这个问题。
该学说以为:
在催化某一反映时候,酶一方面与底物形成不稳定中间产物,然后中间产物再分解,释放出酶及产生反映产物。
可用公式表达为:
这样,把本来无酶参加一步反映
提成了两步进行。
这两步反映所需要活化能比本来一步反映低,从而加快反映速度。
显然,酶之因此减少反映活化能是由于酶与底物生成了中间产物从而变化了反映途径所致。
3.“钥匙-锁”学说和“诱导契合”学说
酶和底物是如何结合成中间产物?
又如何完毕其催化作用?
1890年。
E.Fischer提出“钥匙-锁”学说,以为酶和底物结合时,底物构造必要和酶活性部位构造非常吻合,就像锁和钥匙同样,这样才干紧密结合形成中间产物。
这在一定限度上解释了酶促反映特性,如专一性;但该学说把酶构造当作是固定不变,这是不切实际,并且该模型不能解释可逆反映。
1958年,D.E.Koshland提出了“诱导契合”学说,克服了“钥匙-锁”模型缺陷,以为酶与底物结合时,底物能诱导酶分子构象变化,使酶能与分子较好地结合,从而发生催化作用。
如图1-1-7所示。
4.使酶具备高催化效率因素
酶为什么比普通催化剂具备更高催化效率?
重要有如下因素:
(1)邻近定向效应指底物和酶活性部位邻近,对于双分子反映来说也包括酶活性部位上底物分子之间接近,而互相接近底物分子之间,以及底物分子与酶活性部位基团之间还要有严格定向(对的立体化学排列)。
这样就大大提高了活性部位上底物有效浓度,使分子间反映近似于分子内反映,同步还为分子轨道交叉提供了有利条件;使底物进行反映活化能减少,从而大大地增长了酶-底物中间产物进入过渡态几率。
(2)“张力”和”形变”底物结合可以诱导酶分子构象变化,而变化酶分子又使底物分子敏感键产生“张力”甚至“形变”,从而增进酶-底物中,间产物进入过渡态。
(3)酸碱催化酶活性部位上某些基团可以作为良好质子供体或受体对底物进行酸碱催化。
(4)共价催化某些酶可以和底物生成不稳定共价中间物,这种共价中间物进一步生成产物要比非催化反映容易得多。
(三)影响酶催化反映因素
1.酶浓度影响
在酶促反映中,如果底物浓度大到足以使酶饱和,则反映速度与酶浓度成正比(图1-1-8):
V=k[E]。
2.底物浓度影响
(1)底物浓度对酶促反映速度影响
在酶浓度等条件恒定,反映系统中没有不利于酶发挥作用因素存在时,用反映速度对底物浓度作图得始终角双曲线(图1-1-9)。
由曲线可以看出:
当底物浓度[S]较低时,反映速度和底物浓度几乎成正比。
当底物浓度较高时,反映速度也随浓度增长而升高,但不明显。
当浓度大大增长时,反映速度趋近一种最大值即最大速度Vmax,此时反映速度与底物浓度无关。
3.PH值影响
酶经常限于某一pH值范畴内才体现出最大活力,这种体现出酶最大活力pH值就是酶最适pH值。
当pH高于或低于这个最适值时,酶活性就会减少。
普通典型最适pH曲线为钟型曲线(图1-1-10)。
pH值对酶活性影响因素,除了由于过酸或过碱使酶变性失活外,重要是由于影响了酶分子活性中心上关于基团解离或底物解离,这样就影响了酶与底物结合,从而影响了酶活力。
4,温度影响
各种酶在一定条件下均有一种最适温度,在最适温度两侧,反映速度都较低,呈钟罩形曲线(图1-l-11)。
温度对酶促反映影响有两个方面:
一方面是温度升高,反映速度加快,与普通化学反映相似;另一方面,随着温度升高,酶蛋白变性也随之增长,减少有活性酶数量,减少了酶促反映速度。
酶促反映最适温度就是两种过程平衡。
在低于最适温度时,前一种效应为主,在高于最适温度时,后一种效应为主。
5,激活剂影响
能提高酶活性物质称为激活剂。
按分子大小可分为3类:
第一类为无机离子,如Mg2+是各种激酶激活剂,C1-能激活唾液α-淀粉酶;第二类为中档大小有机化合物,一种是还原剂,如半胱氨酸、还原型谷胱甘肽等,另一种是金属螯合剂,能除去酶中重金属杂质,从而解除重金属对酶抑制,如乙二氨四乙酸(EDTA);第三类为蛋白质性大分子化合物,此类激活剂用于酶原激活,使无活性酶原变成有活性酶。
6.抑制剂影响
某些物质,不引起酶蛋白变性,但能使酶分子上某些必须基团发生变化,因而引起酶活性下降,甚至丧失。
这种作用称为抑制作用,起抑制作用物质称为抑制剂。
酶抑制作用分为不可逆抑制作用和可逆抑制作用两类。
(四)酶分类和命名
1.酶国际系统分类法
国际生物化学联合会酶学委员会提出酶国际系统分类法分类原则是:
将所有已知酶按其催化反映类型
 升级会员
升级会员 