整理淀粉酶类的生产Word格式.docx
《整理淀粉酶类的生产Word格式.docx》由会员分享,可在线阅读,更多相关《整理淀粉酶类的生产Word格式.docx(14页珍藏版)》请在冰豆网上搜索。
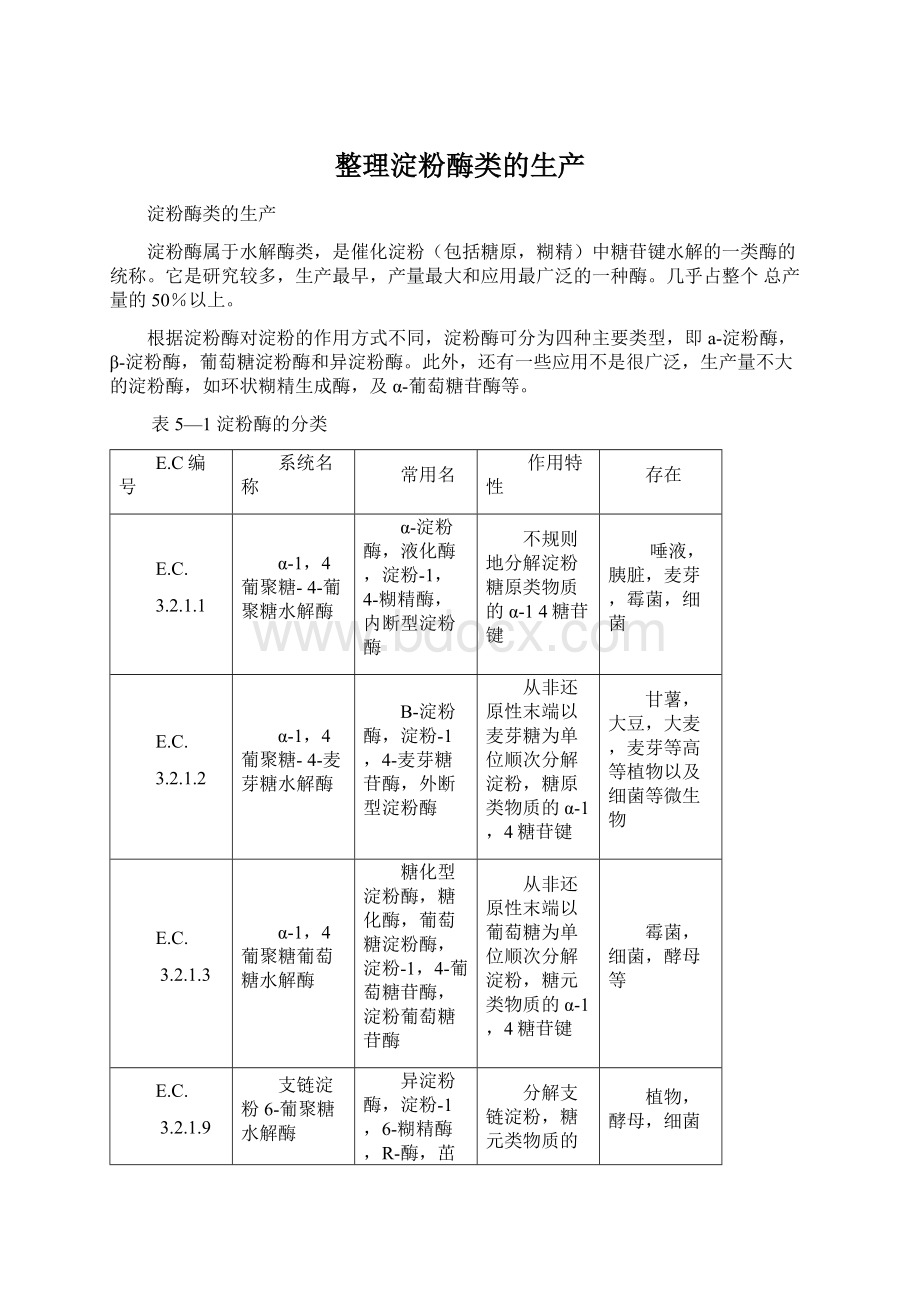
从非还原性末端以葡萄糖为单位顺次分解淀粉,糖元类物质的α-1,4糖苷键
霉菌,细菌,酵母等
3.2.1.9
支链淀粉6-葡聚糖水解酶
异淀粉酶,淀粉-1,6-糊精酶,R-酶,茁酶多糖酶,脱支酶
分解支链淀粉,糖元类物质的α-1,6糖苷键
植物,酵母,细菌
淀粉酶的种类不同,对直链淀粉和支链淀粉的作用方式也不一样。
各种不同的淀粉酶对淀粉的作用有各自的专一性。
淀粉是自然界中分布极广的碳水化合物,它是由葡萄糖基相连接聚合而成的,根据连接方式不同一般可将其分为直链淀粉和支链淀粉两种。
直链淀粉的葡萄糖基几乎都是以α-1,4键相互连接成的直连,聚合度为100—6000个葡萄糖单位不等,最近研究认为直链淀粉分子中也有极少量的分枝结构存在。
支链淀粉则较复杂,除有较多的α-1,4键连接外,还在分子内有α-1,6键连接成树枝状,聚合度也比直链淀粉高。
表5—2常见淀粉中直链与支链淀粉含量
淀粉品种直链淀粉/%支链淀粉/%
玉米2773
马铃薯2377
甘薯2080
木薯1783
大米1783
糯玉米0100
糯高粱0100
糯米0100
5.1α-淀粉酶的生产
α-淀粉酶作用于淀粉时,可以随机的方式从分子内部切开α-1,4葡萄糖苷键而生成糊精和还原糖。
其水解位于中间的α-1,4键的概率比水解位于分子末端的概率大,不能水解支链淀粉的α-1,6键,也不能水街紧靠1,6分支点的-α-1,4键,不能水解麦芽糖,但可以水解含有3个或3个以上α-1,4糖苷键的低聚糖。
由于水解产物的还原性末端葡萄糖残基C1碳原子为α构型,故称α-淀粉酶。
目前,国内外生产α-淀粉酶所采用的菌种主要有细菌和霉菌两大类,典型的与芽孢杆菌和米曲霉。
米曲霉常用固态曲法培养,其产品主要用作消化剂,产量较小,芽孢杆菌则主要采用液体深层通风培养法大规模地生产α-淀粉酶,如我国的枯草杆菌BF—7658.
5.1.1α-淀粉酶的性质
几种微生物α-淀粉酶的性质见表5—3
表5—3各种α-淀粉酶的性质
作用机制
酶来源———————————耐热性/℃pH稳定性适宜Ca2+的保护作用
淀粉分解限度/%主要水解产物(15min)(30℃,24hpH
枯草杆菌糊精,麦芽糖
(液化型)35(30%)葡萄糖6%65~804.8~10.65.4~6.0+
枯草杆菌
(糖化型)70葡萄糖(41%)
麦芽糖(58%)
麦芽三糖,糊精55~704.0~9.04.8~5.2—
(耐热型)35糊精,麦芽糖,
葡萄糖75~905.0+
米曲霉48麦芽糖(50%)55~704.7~9.54.9~5.2+
黑曲霉48麦芽糖(50%)55~704.7~9.54.9~5.2+
黑曲霉
(耐酸性)麦芽糖(50%)55~701.8~6.54.0+
根霉48麦芽糖(50%50~605.4~7.03.6—
———————————————————————————————————————
1.pH对酶活性的影响
一般α-淀粉酶在pH5.5~8稳定,pH4以下易失活,酶活性的最适pH5~6,即在此pH条件下酶的催化反应速度最快,另外酶的催化活性和酶的稳定性是有区别的,前者指酶催化反应速度的快慢,活性高反应速度快,反之则反应速度慢,而后者表示酶具有催化活性而不失活。
酶最稳定的pH不一定是酶活性的最适pH,反之,酶活性的最适pH不一定使酶最稳定。
在霉菌中,黑曲菌α-淀粉酶耐酸性强,黑曲霉NRRL330α-淀粉酶的最适pH为4.0,在pH2.5,40℃处理30min尚不失活,然而在pH7.0时,55℃处理15min,几乎没有损失,而在pH2.5处理则完全失活。
枯草杆菌α-淀粉酶作用的最适pH为5~7.嗜碱细菌中存在着最适pH为4.0~11.0的α-淀粉酶。
嗜碱性芽孢杆菌NRRLB3881α-淀粉酶的最适pH9.2~10.5,嗜碱性假单胞杆菌α-淀粉酶的最适pH为10.。
各种不同的酶的最适pH可以通过实验测定,由于最适pH受底物种类。
浓度,缓冲液成分,温度和时间等因素的影响,测定时必须控制一定的条件,条件可以改变可能会影响最适pH。
2.温度对酶活性的影响
温度对酶活性有很大的影响。
纯化的α-淀粉酶在50℃以上容易失活,但是有大量Ca2+存在下,酶的热稳定性增加。
芽孢杆菌的α-淀粉酶耐热性增加。
芽孢杆菌的α-淀粉酶耐热性较强。
枯草杆菌α-淀粉酶在65℃稳定。
嗜热脂肪芽孢杆菌和凝结芽孢杆菌的α-淀粉酶的热稳定性更强,前者经85℃处理20min,尚残存酶活70%,后者在Ca2+存在下,90℃时的半衰期长达90min。
有点嗜热芽孢杆菌的α-淀粉酶在110℃仍能液化淀粉。
地衣芽孢杆菌的α-淀粉酶其热稳定性不依赖Ca2+,可在EDTA存在下测定酶活,以区别于非耐热性α-淀粉酶。
霉菌α-淀粉的耐热性较低,黑曲霉耐酸性α-淀粉酶的耐热性比其非耐热性α-淀粉酶为高,在pH4,55℃加热24h也不失活。
然而拟内孢霉α-淀粉酶在40℃以下也很不稳定。
α-淀粉酶在各种酶中是耐热性较好的酶,其耐热程度一般是按动物α-淀粉酶,麦芽α-淀粉酶,丝状菌α-淀粉酶,细菌α-淀粉酶的顺序而增强。
曾对各种α-淀粉酶粗制剂的水溶液进行加热处理,每分钟升高1.5℃,直至80℃,发现各种酶的残留活性是:
真菌来源的为1%,谷物来源的为25%,细菌来源的为92%。
α淀粉酶的耐热性还受底物的影响,在高浓度的淀粉浆中,最适温度为70℃的枯草杆菌α-淀粉酶,在85—90℃时的活性最高。
3.钙与α-淀粉酶活性的关系
α-淀粉酶是单成分酶,大多数α-淀粉酶活性需要钙离子,钙离子对酶的稳定性起重要作用。
Ca2+使酶分子保持适当的构象,从而维持其最大的活性与稳定性。
钙和酶的结合牢度依次是:
霉菌>细菌>哺乳动物>植物。
Ca2+对麦芽α-淀粉酶的保护作用最明显。
枯草杆菌糖化型α-淀粉酶(BSA)同Ca2+的结合比液化型(BLA)更为紧密。
向BSA中添加Ca2+对酶活性几乎不发生影响,单用EDTA处理也不能引起失活,只有在低pH(pH3.0)下用EDTA处理才能去除Ca2+,但若添加与EDTA当量的Ca2+,并将pH恢复至中性,则仍然可恢复它的活性。
除Ca2+外,其他二价碱土金属Sr2+,Ba2+,Mg2+等也有使无Ca2+的α-淀粉酶恢复活性的能力。
枯草杆菌液化型α-淀粉酶(BLA)的耐热性因Ma+,C1-和底物淀粉的存在而提高,NaC1与Ca2+共存时对提高α-淀粉酶的耐热性的作用尤为显著。
添加Ca2+有助于增加酶的热稳定性,但实际上淀粉中所含微量Ca2+已足够酶的充分活化所需。
5.1.3α-淀粉酶对底物的水解作用
1.α-淀粉酶的水解方式
电费是由葡萄糖单位组成的大分子。
它与水在催化剂的作用下生成较小的糊精,低聚糖,直至最小构成单位——葡萄糖,这个过程称为淀粉的水解。
淀粉的水解可用酸或淀粉酶作为催化剂。
酶水解具有较强的专一性,不同的酶作用于不同的键,如α-淀粉酶从淀粉分子内部随机切割α-1,4键,但不能水解α-1,6键,α-1,3键,甚至不能水解紧靠分支点的α-1,4键。
同时,酶催化反应具有条件温和,设备简单,副反应极少等优点。
而酸水解没有专一性,同时可以水解α-1,4键,α-1,6键及α-1,3键等。
另外,淀粉通过水解反应的葡萄糖,受酸和热的作用,一部分又发生复合反应和水解反应,影响葡萄糖的产率,增加糖化液精致的苦难。
α-淀粉酶对于直链淀粉的作用第一步是将直链淀粉任意的迅速降解成小分子糊精,麦芽糖和麦芽三糖,第二部缓慢地将第一步生成的低聚糖水解为葡萄糖和麦芽糖。
由于α-淀粉酶不能切开支链淀粉分支点的α-1,6键,也不能切开α-1,6键附近的α-1,4键,但能越过分支点而切开内部的α-1,4键,因此水解产物中出了含有葡萄糖,麦芽糖意外,还残留一系列具有α-1,6键的极限糊精,和含4个或者更多葡萄糖残基的带α-1,6键的低聚糖。
表5-5是枯草杆菌α-淀粉的水解产物分布,表中数据位不同聚合度的低聚糖占总糖分的百分率,不同来源的α-淀粉酶,水解产物存在差别。
表5-5枯草杆菌α-淀粉酶的水解淀粉产物分布单位:
%
直链淀粉支链淀粉
水解产物——————————————————————————————
60min180min60min180min
G12.35.31.43.3
G210.112.35.58.3
G312.822.08.210.8
G46.010.50.92.5
G510.214.84.96.7
G620.630.114.026.8
G714.75.19.89.2
高分子物质23.3055.332.4
注:
G1,G2…..表示葡萄糖聚合度。
2.α-淀粉酶的水解极限
当α-淀粉酶作用于淀粉时,随着反应的进行,溶液黏度逐渐下降而还原力逐渐增加。
由于底物浓度减少,产物浓度增加,酶可能部分失活,最后导致反应速度降低,直至还原力不在增加,此时的水解率称为“水解极限”。
不同来源的α-淀粉酶,水解极限各部相同,一般α-淀粉酶水解率为40%~50%,但黑曲霉ATCC15475的水解率可达95%~100%,拟内孢霉α-淀粉酶水解率可达90%,其产物均是葡萄糖。
枯草杆菌糖化型α淀粉酶作用于可溶性淀粉时,水解率达70%以上,而淀粉液化芽孢杆菌所产液化型α-淀粉酶的水解率只有30%。
假定直链淀粉被彻底水解,即水解极限为100%,则成成13份葡萄糖及87份麦芽糖,而当具有4%分支的支链淀粉被彻底水解。
则生成73份麦芽糖,19份葡萄糖和8份异麦芽糖。
5.1.4α-淀粉酶的工业生产
1.菌种
工业上大规模生产和应用的α-淀粉酶主要来自细菌和曲霉。
芽孢杆菌所产α-淀粉酶分为液化型与糖化型两种。
目前只有液化型酶有用,由于活性高,发酵周期短,酶的耐热性高,尤其是枯草杆菌为大多数工厂所采用。
我国淀粉糖工业使用的液化酶BF-7658,美国的Tenase等属于这一种。
地衣芽孢杆菌的酶耐热性比枯草杆菌为高,但产量较低。
芽孢杆菌易于退化和遭受噬菌体感染而降低产酶能力。
由微生物制备酶制剂,产酶量高,易于分离和精制,适于大量生产。
当然亦能从植物和动物中提取α-淀粉酶,满足特殊的需要,但由于成本高,产量低,目前还不能实现工业化生产。
具有使用价值的α-淀粉酶生产菌列于表5-6.
表5-6常用的α-淀粉酶生产菌
枯草杆菌JD-32马铃薯杆菌
枯草杆菌BF-7658嗜热糖化芽孢杆菌
淀粉液化芽孢杆菌多黏芽孢杆菌
嗜热脂肪芽孢杆菌嗜碱假单胞菌
嗜热硬脂芽孢杆菌溶淀粉变种黑曲霉
淀粉糖化芽孢杆菌米曲霉
地衣形芽孢杆菌泡盛酒曲酶
由于不断的选育改良,现在工业生产上用的菌种产生α-淀粉酶的能力已是原始菌株的数倍乃至数十倍,例如淀粉液化芽孢杆菌ATCC23844的α-淀粉酶活性,每1mL已达456000U,地衣芽孢杆菌ATCC9789,用y射线,NTG,之外先单独或交叉处理7次后,其耐热性α-淀粉酶活性增加25倍,结合培养条件的改进而用于工业生产。
紫外线处理肉桂色曲霉,耐酸性α-淀粉酶活性提高了6倍。
我国生产菌株枯草杆菌BF7658-209,是由野生型菌株,经物理和化学方法交叉处理得到的变异株,其产酶活性比母株高5.0%。
此外,用X射线也曾得到高产突变株。
连续使用同一诱变剂时,由于发生平顶效应(Plateaueffect),诱变效果会随着处理次数而降低。
此时必须更换诱变手段。
在使用紫外线,y射线,快中子等为诱变剂处理米曲霉时,高剂量不一定有不利于高产变株的生成。
近年来,利用转化法改良菌株,在枯草杆菌α-淀粉酶的生产菌上已取得可喜进展,例如α-淀粉酶活性高而耐热性差的枯草杆菌纳豆株的DNA转入耐热性强而酶活低的枯草杆菌Marburg株中,结果引起了后者遗传性状的改变,其α-淀粉酶提高了14倍,蛋白酶活性提高了5倍,这个杂交种所产酶也具有亲株性能。
但由于酶活性还不及生产菌株而尚未实用。
又将生产菌的DNA转入枯草杆菌Marburg株,得到酶活性较高的转化株SP-38,又将SP-38DNA转入Marburg使酶活性有了进一步提高。
不同生态环境下分离的微生物,它的α-淀粉酶性质与其生长环境相适应,从温泉分离的一株耐热解淀粉假单胞杆菌,它的α-淀粉酶最适作用温度65~80℃。
在55℃培养的嗜热脂肪芽孢杆菌和凝结芽孢杆菌,所产的α-淀粉酶在90℃下几乎不失活。
分别在35℃与55℃培养凝结芽孢杆菌,所产α-淀粉酶的热稳定性55℃与43℃下培养嗜热脂肪芽孢杆菌,其α-淀粉酶的各种理化性质几乎无明显区别,此外,地衣芽孢杆菌虽在30℃培养,它的α-淀粉酶最适温度80~85℃,耐热性很强。
生产概要
霉菌的α-淀粉酶大多采用固体曲法生产,细菌α-淀粉酶则以液体深层发酵为主。
固体培养以麸皮为主要原料。
酌量添加米糠或豆饼的碱水浸出液,以补充氮源。
在相对湿度90%以上,芽孢杆菌用37℃,曲霉用32~35℃培养36~48h后,立即在40℃下烘干或风干,即成工业生产用的粗酶。
液体培养常以麸皮,玉米粉,豆饼粉,米糠,玉米浆等为原料,并适当补充硫酸铵,氯化铵,磷酸铵等无机氮源,此外还需添加少量镁盐,磷酸盐,钙盐等。
固形物浓度一般为5%~6%,高者达15%,为了降低培养液黏度,淀粉原料可用α-淀粉酶液化,氮源可用豆饼碱水浸出液代替。
以霉菌为生产菌时,宜采用微酸性,而细菌宜在中性至微碱性培养,培养温度霉菌32℃,细菌37℃,通风搅拌培养时间24~48h。
当酶活达到高峰时结束发酵,离心或以硅藻土作助滤剂滤去菌体及不溶物。
在Ca2+存在下低温真空浓缩后,加入防腐剂(松油,麝香草粉,苯甲酸钠等),稳定剂(5%~15%食盐和钙盐,锌盐或山梨醇)以及缓冲剂后就成为成品。
为了提高它的耐热性,也可在成品中添加少来硼酸盐。
这种液体的细菌α-淀粉酶呈暗褐色,带不快之臭味,在室温下可放置数月而不失活。
为了制备高活性的α-淀粉酶,并使贮运方便,可用硫酸铵盐析或溶剂沉淀制成粉状酶制剂。
在由Ca2+存在下降浓缩发酵液调节pH到6左右,加入40%左右硫酸铵静置沉淀,倾去大部分清液后,加入硅藻土助滤剂,收集沉淀于40℃以下风干,为了加速干燥,减少失活,酶泥中可拌入大量硫酸钠,磨粉后加入淀粉,乳糖,CaCl2等作稳定填充剂后即成为成品。
麸曲可以用水抽提出进行盐析。
为了减少色素的溶出,麸曲必须进行风干抽提液中如色素较多,可用CaCl2和NaHPO4形成不溶性沉淀而吸附除去。
溶剂(酒精,丙酮等)沉淀时,为减少酶的变性,宜在低温下(15℃左右)操作,在由CaC
L2,乳糖,糊精等存在下,加入冷却的溶剂至最终浓度70%,收集沉淀用无水酒精脱水,40℃以下烘干。
有些菌株产生一定比例的蛋白酶,这样不但妨碍使用效果,还会引起α-淀粉酶在贮藏过程中失活,夹杂的蛋白酶量越大,失活就越严重。
培养基中添加柠檬酸盐可抑制某些菌株产生蛋白酶,枯草杆菌发酵液中伴生的蛋白酶可可借加热50-60℃处理而消除。
此外细菌α-淀粉酶尚可利用底物淀粉吸附,而同蛋白酶分开。
为了提高淀粉的吸附效果,淀粉可以膨胀处理。
淀粉吸附法的主要步骤如下:
调节酶液的pH到6.0,加18%硫酸铵搅匀,并以玉米淀粉与硅藻土的5:
1的混合物分散在18%硫酸铵溶液中,倒于漏斗上形成底层,再讲上述酶液通过滤层用15%硫酸铵液洗涤后,用含0.001mol/LCaCl2的0.04mol/L磷酸缓冲液洗脱,经DuoliteA-2树脂脱色,再用40%硫酸铵盐析或60%丙酮沉淀,这样可以制备纯度极高的产品。
3,影响α-淀粉酶生产的因素
1)固体培养
以麸皮为主要原料时,添加少量豆饼或豆饼的碱水浸出液等有机氮源对产酶有益。
原料洒水以1:
1:
2为宜。
枯草杆菌要求洒水率比霉菌稍高。
固体培养时,培养基的碳氮比生产酶影响不如液体培养时明显,培养枯草杆菌时,培养基的初pH以杀菌后6.3~6.4为宜。
如果适当添加米糠,保持初pH6.0~6.5可使产酶稳定。
生产α-淀粉酶的最适温度范围比较小,在整个培养过程中,品温不能有7~8℃只差。
最适温度为37℃的枯草杆菌,品温达45℃时,产酶就降低。
2)液体深层培养
(1)碳源的诱导及阻遏微生物生产的α-淀粉酶可以说是半组成酶,因为大多数工业生产的淀粉酶菌种,例如淀粉也化芽孢杆菌,枯草杆菌168,地衣形芽孢杆菌以及米曲霉等,即使培养基中不含淀粉或者不含具有α-1,4键的多糖或低聚糖,仍然可以生成α-淀粉酶,但是它们的产量可收到淀粉或其他α-1,4麦芽寡糖的诱导而增加。
(2)福本指出,在液体静止培养下各种碳源对淀粉也化芽孢杆菌的效果依次是:
可溶性淀粉>麦芽糖>甘露醇>阿拉伯糖>葡萄糖>蔗糖>乳糖>半乳糖>木糖。
对枯草杆菌BF-7658研究,得到类似结果(表5-7)。
豆饼浸出液为氮源时,淀粉液化芽孢杆菌的最佳碳源依次是乳糖,半乳糖,淀粉,麦芽糖。
用在葡萄糖与氨构成的合成培养基中,乳糖仍最适合于菌体生长,糊精则完全无效,但琥珀酸,延胡索酸几乎与可溶性淀粉一样有效,葡萄糖,果糖,蔗糖等在高浓度时抑制α——淀粉酶的生成。
表5-7枯草杆菌BF-7658对各种碳源的利用
碳源终pH酶活性/(U/ml)初浓度/%残糖量/%糖利用率/%
可溶性段百分比(C.P.)7.5413.881.5659.7
糊精(实验试剂)7.5384.162.3144.3
蔗糖(C.P。
)7.011
麦芽糖(C.P.)7.5443.750.9175.8
乳糖(C.P.)7.0213.651.3762.1
半乳糖(C.P.)7.019
葡萄糖(A.P.)7.0143.560.9772.8
木糖(C.P.)4.134.0
-*容易利用的碳源,例如葡萄糖,果糖,蔗糖等只能促进细胞的呼吸与生长,而不利用α-淀粉酶生产,代谢愈快的糖对α-淀粉酶生产的抑制越严重,但是一些作为能源利用性很差的糖类,像糖原,乳糖,半乳糖,棉子糖等却能促进α-淀粉酶的合成,乳糖对芽孢杆菌NRRLB3881生产碱性淀粉酶却不适合。
麦芽糖与淀粉,糖原一样对米曲霉生产α淀粉酶有促进作用,麦芽四糖对嗜热脂肪芽孢杆菌,地衣形芽孢杆菌,异麦芽糖对米曲霉,麦芽糖对嗜热芽孢杆菌碱性α-淀粉酶生产的诱导作用最强。
碳源种类与α淀粉酶及胞内核糖核酸的形成有着密切关系。
当核糖与淀粉共存时,α-淀粉酶的产量比单独用淀粉时高一倍。
葡萄糖等易利用碳源,在浓度高时,妨碍α-淀粉酶的生成,这是一种分解代谢物阻遏。
它在α-淀粉酶的生物合成上起着一种调节作用。
Schaeffer(1969)认为枯草杆菌的α-淀粉酶生产受到分解大写唔阻遏的控制。
Mers等(1972)报道地衣芽孢杆菌α-淀粉酶的生产可受到葡萄糖或其他低相对分子质量可代谢糖的抑制,但是Coleman报道,淀粉液化芽孢杆菌生长培养物中添加葡萄糖,并不引起对淀粉酶合成的分解代谢阻遏。
Meyrath等报道,葡萄糖等易利用的糖,在浓度高时,对米曲霉α-淀粉酶的阻遏作用只发生在生长的早期。
为了提高α-淀粉酶的产量,避免分解代谢物的阻遏,除选育康分解代谢的变异菌株外,就是采用代谢慢慢的碳源,例如乳糖,乳清。
但大多数工厂使用淀粉为碳源,这不仅可避免分解代谢物阻遏,还具有诱导作用。
为了降低培养基的黏度,杀菌时旺旺添加α-淀粉酶液化,在液化过程中生成的葡萄糖,仍有造成阻遏的可能,因此将碳源采取流加法加入,是促进产酶的好办法。
(2-)氮源福本以淀粉为氮源,培养液化淀粉芽孢杆菌时,白蛋白,酪蛋白,大豆饼碱水抽提液,据蛋白胨为较优氮源。
玉米浆与其他蛋白质并用时也是良好氮源。
以酪蛋白的酸或碱水解物作氮源时,产酶不及酪蛋白本身。
各种氨基酸与酪蛋白的酶水解物有利于α-淀粉酶的生成,谷氨酸促进黑曲霉α-淀粉酶的生成。
氮源时细胞合成α-淀粉酶的原料,香淀粉液化芽孢杆菌洗涤细胞用时添加碳源与氨基酸时,一些氨基酸比氨的效果好,一些氨基酸则逊于氨,另一些氨基酸则起抑制作用,混合氨基酸的效果比单一氨基酸好,天冬酰氨对生产α-淀粉酶有效,而甘氨酸由抑制。
甘氨酸的抑制作用能用丙氨酸,蛋氨酸与谷氨酸接触。
酪蛋白,豆饼的热,碱水抽出物是工业上最优良的氮源。
基槽废液是黑曲霉NRRL330α-淀粉酶生产的一种最好的氮源,以5%玉米粉加7°
Be酒糟水,α-淀粉酶产量最高。
硝酸钠,硫酸铵,氯化铵,硝酸铵,醋酸铵及尿素等无机氮均可在不同程度上增加α-淀粉酶的产量,但大多喜用硝酸钠。
黑曲霉NRRL330以8%的甘薯为碳源,并添加0.3%NaNO3,α-淀粉酶活性可增加10倍。
用Minoda培养基(淀粉2%,麸皮4%,(NH4)2SO40.2%,KH2PO40.1%,CaCO32.0%)培养黑曲霉时,其耐酸性α-淀粉酶较用其他培养基时高2倍。
若在添加NaNO3,则酶活性可增加3倍。
NH4NO3与NH4Cl对黄曲霉生产α-淀粉酶的效果比有机氮好,NH4NO3亦是米曲霉的最佳氮源。
培养液pH应保持在最适pH范围。
用米曲霉生产淀粉酶时,以硫酸铵为氮源。
用米曲霉生产淀粉酶时,以硫酸铵为氮源,若同时添加醋酸钠,则其浓度虽高达1.6%,发酵液的pH仍能维持在最适状态。
第1页(3)碳氮比黑曲霉NRRL330生物合成α-淀粉酶与糖化酶时,碳氮比与产煤量之间并无一定规律。
且培养液的终pH也同碳氮比无关。
以玉米与豆饼为碳氮源时,两者比例改变,对于枯草杆菌BF-7658与TUD-127生物合成α-淀粉酶由明显影响,总固形物13%~14%。
以上,玉米与豆饼之比以(8~9):
5为宜。
安全预评价方法可分为定性评价方法和定量评价方法。
(4)无机盐α-淀粉酶活性的表现需要Ca2+,但在工业原料本身所含有的Ca2+已能够满足生长与产酶的需要,一般不需另外不加。
发酵中途补加CaCl2稍稍有利于枯草杆菌TUD产酶及残糖分的降低。
添加柠檬酸盐可一直枯草杆菌BS1968产生蛋白酶而不妨碍α-淀粉酶的生产。
直接市场评估法又称常规市场法、物理影响的市场评价法。
它是根据生产率的变动情况来评估环境质量变动所带来影响的方法。
磷酸盐无论对细菌和霉菌的α-淀粉酶的生产都很重要。
在以甘薯8%,硝酸钠0.3%构成的培养基中,培养黑曲霉
 升级会员
升级会员 