整理植物茎的形态与结构.docx
《整理植物茎的形态与结构.docx》由会员分享,可在线阅读,更多相关《整理植物茎的形态与结构.docx(32页珍藏版)》请在冰豆网上搜索。
整理植物茎的形态与结构
第六章 茎的形态与结构
种子萌发后,上胚轴和胚芽向上生长产生茎和叶。
茎端和叶腋内的芽(bud)活动生长,形成分
枝(twig)。
继而新芽不断产生与生长,最后形成了繁茂的植株地上系统。
第一节茎的形态特征
一、茎的发生
茎是植物进化过程中早于根发展起来的营养器官,是植物适应陆生生活、向空中发展的标志性结构之一。
在植物(尤其是种子植物)的个体发育中,茎由胚芽和胚轴发育而来。
在漫长的系统演化进程中,植物的茎沿着从草本到木本、从木本到草木本共荣的方向发展。
最初植物的茎是多细胞束状聚集的、无组织分化、由外而内不能分清结构层次的“假茎”;随着生存质量的改善和环境的选择作用,组织进化和发展起来;于是,不同组织间的分工协作,使真正意义上的茎得以进化形成,植物便能更好地利用环境、适应环境、保护和繁衍自身。
苔藓植物、蕨类植物、裸子植物和被子植物相应地应运而生、进化出现。
由于木本植物的茎可塑性小,适应力差,生活周期长,结实率相对较低,所以,古生代的鳞木(Lepidodendron)、封印木(Sigillaria)等高大的乔木,到二叠纪全部灭亡。
现存的木本植物的数量远少于草本植物的数量。
草本植物的进化,从多年生到一年生。
因为,草本植物的茎生活周期短,生殖阶段发生早,仅用极少限量的物质消耗在营养器官的构建上,而种子的繁殖却达到了最大限度。
种子繁衍代数多,变异几率大,在地质年代的气候变迁和冰川袭击中,草本植物则适应而生,显示了更强的适应性。
因此,茎的出现是植物进化、繁荣过程中的又一个大事件。
茎的发育,使植株上的芽、叶等有更大的发展空间。
茎中形成的维管组织可以长距离地输送水分和养分,更加促进了茎的生长和发育,形成发达的茎枝系统,于是更多的芽、叶甚至被子植物的花等器官孕育产生。
多样性生存环境的长期作用和影响,使植物的茎更是千姿百态。
植株上数目繁多的叶在一定的空间呈镶嵌分布,充分地接受光照和进行光合作用制造营养物质。
同时,茎枝系统又支持着大量的花和果实,使它们更适合发育生长,更有利于传粉及果实和种子的生长、传播,繁殖后代。
这是植物对陆生环境的有效适应。
二、茎的生理功能和应用
(一)茎的生理功能(茎是植物的营养器官之一,其生理功能主要是支持和输导作用。
有些植物的茎还由于其部分或全部地特化为变态器官,从而具有繁殖、攀援、保护等特殊功能。
1.支持作用
大多数被子植物的主茎直立生长于地面,分生出许多大小不同的枝条,并着生数目繁多的叶,主茎和枝统称为茎。
茎支持植株上分布的叶、花和果实,使它们彼此镶嵌分布,更利于光合作用和果实、种子的发育与传播。
2.输导作用
茎连接着植株的根和叶,根部从土壤中吸收的水分、矿质元素以及在根中合成或储藏的有机营养物质,要通过茎输送到地上各部;叶进行光合作用所制造的有机物,也要通过茎输送到体内各部被利用或储藏。
因此,茎是植物体内物质输导的主要通道。
3.储藏、繁殖和光合作用
茎还有储藏、繁殖和光合作用等功能。
有些植物可以形成鳞茎、块茎、球茎、根状茎等变态茎,储藏大量养分,并可以进行营养繁殖。
还可利用某些植物的茎易产生不定根和不定芽的特性,采用枝条扦插、压条和嫁接等方法繁殖植物。
此外,一些植物的叶却退化或早落,茎呈绿色扁平状,可终生进行光合作用,如假叶树(Ruscusaculeatus)、竹节蓼(Homalocladiumplatycladum)、昙花(Epiphyllumoxypetalum)、仙人掌(Coryphanthaelephantidens)等的绿色肉质茎。
有的茎中还有大量的大型薄壁组织,富含水分,而发展成为储水组织。
还有一些植物,如石榴、山楂、皂荚,茎的分枝变为刺,具保护作用。
葡萄、南瓜等植物的茎的分枝还会变为卷须,成为重要的攀援器官而具有攀援作用(详见第八章)。
)
(二)茎的利用(植物的茎常因特殊的结构组成、积累储藏某些特定的代谢物质等,被人们广泛利用。
茎的利用通常有以下几个方面:
1.工业原料
(1)材用:
各类木本植物的茎因其木质化程度很高,作为木材被广泛用于建筑、桥梁、家具、工艺雕刻等多种行业领域。
如松、杉、柏、楠木、榆树等,以及软木、竹材等。
(2)编织:
有些植物的茎较为柔韧,可作编织的原料,如省藤[Daemonoropsmargritae(Hance)Becc]、框柳(Salixhoriganagikimura)等。
(3)纤维:
有些植物茎的韧皮部中含有发达的纤维,可以用作纺织、麻绳、麻袋等的原料。
如苎麻、黄麻、亚麻等。
(4)化工原料:
有些植物的茎中含有特殊物质,而可通过多种方法,加以提取利用。
如橡胶、生漆、树胶、糖料、淀粉等均可从植物体忠提取获得。
2.食用、药用
有些植物的茎主要由含大量营养物质的薄壁细胞组成,可以食用,如莴苣(LactuceindicaL.)的肉质茎、马铃薯的块茎等。
有些植物的茎中积累的代谢产物因具有一定的药用价值,可以用作药材,如杜仲(EucommiaulmoidesOliv.)、天麻(GastrodiaelataBlume)等。
3.观赏、艺术品
有些植物的茎由于其形态奇异、色彩斑斓、或经雕琢作为观赏。
如方竹(Chimonobambusaqundrangularis)、紫竹(黑竹)(Phyllostachysnigra)、各种树桩盆景等。
随着科学技术的发展,对茎的利用,特别是综合利用,将会更加广泛。
)
三、茎的形态与组成
(一)茎的形态
植物的茎是在复杂的地理、气候环境中进化形成的。
植物的茎一般包括主茎(干)和各级分枝两部分。
不同植物,其茎的形态特征不同。
多数植物的茎呈辐射对称的圆柱体;有些植物的茎呈三棱形,如莎草科植物;或四棱形,如唇形科植物薄荷、留兰香等;或多棱形,如芹菜等。
多数植物的茎实心,如棉花、玉米等;也有一些植物的茎有髓腔因而空心,如禾本科的毛竹、小麦等。
茎的大小因种和环境而异。
有的矮小、幼嫩、可直立生长;有的高大、挺立、不断增粗并高度木质化;有的柔弱不能直立,或攀援或缠绕、或贴附于其他物体,蔓延生长(有关茎的性质,详细参见第三篇第十四章)。
有的茎秆高于100m,如澳大利亚的桉树(Eucalyptusglobulus);而有的植株瘦小,高不过几厘米如牛毛毡,[Eleocharisyokoscensis(Franch.etSavat.)Tangetwang];有的树冠庞大,占地面积可达1500m2以上,如生长于缅甸热带雨林中的榕树(FicusmicrocarpaL.)等。
茎的不同形态都是自身遗传特性决定的,是对环境长期适应的结果。
(二)茎的组成
尽管茎的形态大小和习性千差万变,但其组成基本相似。
一般植物的茎都具有节和节间,节上长叶,茎的顶端或叶腋中有芽。
因此,茎就是枝条上除去叶和芽所留下的轴状部分。
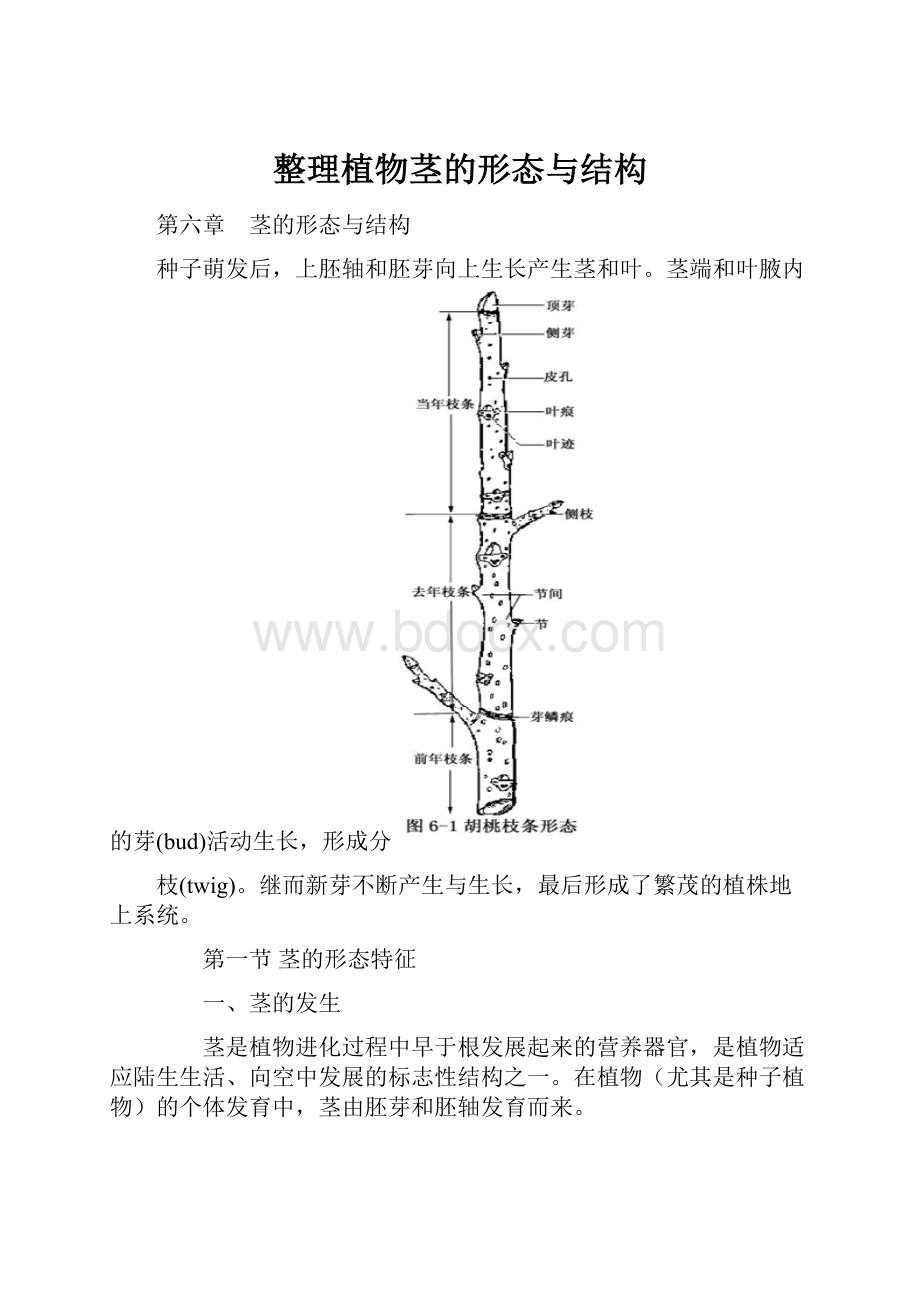
不同植物的节和节间的形状不同,节间的长短亦不同(图6-1)。
不同植物产生的枝条长短不一,同一植株也可以有长枝和短枝之分。
短枝节间短,长枝节间长。
一般长枝是营养生长的枝条(裸子植物例外,如雪松等植物的长、短枝都是营养枝),短枝是开花结实的枝条,又称花枝或果枝。
植株的长、短枝发育是物种的遗传特性决定的。
枝条或节间的长短与植物的种类、枝条发生的位置、年龄、生育期、植株自身的营养状况和环境因素的影响有关。
例如玉米、甘蔗等植株中部的节间较长,茎端的节间较短;水稻、小麦等在幼苗期,各节密集于基部,节间极短,抽穗或抽薹后的节间较长。
有些植物的枝条具有长短枝之分,在果树栽培上,常采取整枝或促控结合的方法来调节长短枝的数量和分布,以获得高产和稳产。
在木本植物的枝条上,其叶片脱落后留下的疤痕称为叶痕(leafscar)。
叶痕中的点状突起是枝条与叶柄间的维管束断离后留下的痕迹,称为维管束迹或称叶迹(folialtrace)。
有的枝条上还有芽鳞痕(budscalescar)存在,这是密集的芽鳞片脱落后留下的环状痕迹。
根据芽鳞痕的特征,可以判断枝条的生长年龄和生长速度(图6-1)。
枝条外表往往可以看见一些小型白色或褐色的斑点称为皮孔(lenticel),这是枝条与外界气体交换的通道。
有些植物的枝条上生长有表皮毛、腺毛等多种类型的毛状附属物,它们或具分泌作用、或具有保护作用。
皮孔、表皮附属物的形态大小、色泽因种而异,是区分物种的参考依据之一。
第二节 芽和茎的分枝方式
一、芽的类型和结构
(一)芽(bud)的类型
芽分布于枝条的顶端或叶腋内,是未发育的枝条或花和花序的原始体。
按照芽生长的位置、性质、结构和生理状态等不同,可将芽分为若干类型。
1.定芽与不定芽(高位芽一般位于地面25cm以上,产生高位芽的植物可分为大高位芽植物(高度超过30米)、中高位芽植物(8~30米乔木)、小高位芽植物(2~8米高的乔木和灌木)、矮高位芽植物(低于2米的灌木),包括乔木、灌木、藤本、高大的草本和附生植物等。
其中每类植物又可分为常绿裸芽、常绿鳞芽和落叶裸芽、落叶鳞芽等次一级类型。
地上芽一般高度为25cm以下,如小灌木、半灌木和小的草本植物。
地上芽贴近地面,生长不利时,地上部分全部或大部分死去,地上部分呈匍匐状或莲座状。
地下芽或隐芽分布于地下或水中,如某些多年生草本、水生或沼生植物等。
研究表明,在温带地区,地下芽和中高位芽对于温度梯度的反应比较灵敏,其次是小高位芽和一年生植物;对于水分梯度的变化,则以地上芽和矮高位芽植物的反应比较强烈和敏感。
了解芽的生活习性,对研究植物的生长与分布、植物在群落中的地位和作用,以及立体种植、人工造林或人造生态群落等均有重要意义。
)
按芽在枝上发生位置是否确定,芽分为定芽(normalbud)和不定芽(adventitiousbud)两类。
定芽在枝上的发生位置固定,顶芽(terminalbud)包括胚芽,只发生于枝的顶端,腋芽(axillarybud)或侧芽(lateralbud)则只发生于叶腋(叶的近轴面与茎的夹角处),顶芽和腋芽都是定芽。
多数植物的一个叶腋中只有一个腋芽,称为单芽(singlebud);有些植物的叶腋可发生2个或几个芽,其中除一个为正芽外,其余均称副芽(accessorybud)(如桃为并生副芽、桂花为叠生副芽)。
法国梧桐等植物的腋芽被包藏于鞘状膨大的叶柄基部内侧,叶柄脱落后腋芽露出,这样的芽称叶柄下芽(infrapetiolarbud)。
不定芽只发生于植株的老茎、根、叶及创伤部位,其发生位置比较广泛,且没有确定性。
如柳的老茎、甘薯的块根、秋海棠的叶上发生的芽都是不定芽。
按芽离地面的高低,芽可分为高位芽、地上芽、地面芽和地下芽或隐芽等类型。
2.叶芽、花芽与混合芽
按结构和性质,芽可分为叶芽、花芽和混合芽。
叶芽(leafbud)是将来发育成营养枝的芽,如水稻的分蘖(侧芽)芽;花芽(flowerbud)是将来发育为花或花序的芽,如广玉兰和小麦的顶芽;混合芽(mixedbud)则为将来同时发育为枝叶和花或花序的芽。
如梨、苹果等植物的顶芽。
叶芽相对瘦小,而花芽和混合芽通常比较肥大,易与叶芽区别。
植物的顶芽和侧芽既可能是叶芽,也可能是花芽或混合芽。
如禾本科植物的顶芽在营养生长期是叶芽,到幼穗分化时则转变成花芽。
植物的副芽则通常可能是花芽,如桃、桂花等的副芽。
3.鳞芽与裸芽
按芽鳞的有无,芽可分为鳞芽和裸芽。
鳞芽(scalybud)是一些生长或起源在冬寒地带的多年生木本植物的芽,有芽鳞片包被,又称被芽。
芽鳞片是叶的变态,其外层细胞角化或栓化、坚硬,外表常被以茸毛、蜡质,或可分泌黏液或树脂,因而可以有效地起到保护作用。
裸芽(nakedbud)是无芽鳞片的仅被幼叶包围着生长锥的芽。
所有一年生、二年生草本和一些多年生木本植物的芽均属于裸芽。
4.活动芽与休眠芽
按芽的生理活动状态,可将芽分为活动芽和休眠芽。
能在当年生长季节中萌发的芽称活动芽(activebud)。
一年生草本植物的芽多数是活动芽。
温带的多年生木本植物,其枝条上近下部的腋芽在生长季节里往往是不活动的,暂时保持休眠状态,这种芽称为休眠芽(dormantbud)。
休眠芽仍具有生长活动的潜势。
在不同的条件下活动芽与休眠芽可以互相转变。
5.珠芽(bulbil)
珠芽是一种未发育的球茎,呈球状、卵圆形等,通常生于叶腋,属于营养繁殖的器官。
如二年生或三年生的半夏[Pinelliaternata(Thunb.)Breit.]叶柄的基部及百合(Lilium.speciosumThunb.)花茎的每一叶柄下部或叶子基部生出的珠芽等。
此外,山药(Dioscoreaspp.)、泽泻科(Alismataceae)、水麦冬科(Juncaginaceae)、天南星科(Araceae)和莎草科(Cyperaceae)部分种类都可在叶柄的基部形成珠芽。
此外,按芽形成的季节分,则生长季中形成并发育的芽称为夏芽,多见于草本和热带常绿植物;在生长季末形成,来年生长季才活动的芽称为冬芽或越冬芽,如多年生植物某些芽。
一个具体的芽,由于分类依据的不同,可给予不同的名称。
如梨的鳞芽可以是顶芽或侧芽,也可以是休眠芽,可以是叶芽,也可以是混合芽。
种子萌发时,胚芽生长,逐渐发育为具叶的枝条。
枝条的顶端和叶腋中生长幼芽。
芽有多种类型,芽在茎或枝条上的位置因植物而不同,芽的位置和活动决定了植物地上部枝条分布的格局。
(二)芽的结构
1.叶芽
叶芽是在植物营养生长的早期陆续出现的芽。
其基本结构包括生长锥(growingtip;growthcone)(芽轴、茎尖分生区)、自内而外体积和发育程度递增的叶原基(leafprimordium)和幼叶(图6-2,图6-3A,B)。
有的植物,芽外方还有一至几片芽鳞,以及从第二、三个(自顶向基)叶原基内侧开始出现的腋芽原基(axillarybudprimordium)。
腋芽原基位于每个叶原基或幼叶的叶腋。
2.花芽
当植株从营养生长转入生殖生长时,开始形成花芽。
花芽是花或花序的原始体,外观较叶芽肥大,内含花或花序各部分(花萼、花瓣、雄蕊和雌蕊)的原基(详见第九章)。
花芽中无叶原基和腋芽原基,茎尖分生组织在花芽分化完成后消失(图6-3C)。
3.混合芽(芽的组成体现了对分生组织主体的保护与对环境的适应。
如密集生长的幼叶或苞片、芽鳞等,它们紧紧地包被着芽轴,使其免遭风吹、雨淋、日晒及一些生物的侵害。
在生长条件不利,或因植物自身发育的需要进入休眠时,芽更是一种绝好的休眠结构——内部细胞进入休眠状态的变化,外部的重重包被和外层栓化的鳞片,限制了氧气注入和水分散失,降低了内部的呼吸与代谢活动。
)
混合芽有完整的叶芽结构,叶原基或幼叶的叶腋内还有花芽原基的分化,混合芽的外围常包被有芽鳞片(图6-3D)。
(三)芽的发生与分枝
植物的分枝源于芽的活动。
因此,了解芽的形成规律是了解植物分枝特征的关键。
关于侧芽的形成有两种观点:
一是侧芽的顶端分生组织即主茎的顶端分生组织(合点起源)由形成叶原基时保留在叶腋处的分生细胞所组成,一是侧芽的顶端分生组织在叶腋内(处)由已分化的细胞恢复分裂能力而形成。
根据拟南芥显性突变体phabulosa(phb)研究表明:
在该类突变体的叶的远轴面出现了侧芽,这就说明侧芽发生于叶的近轴面细胞,而非来自茎端分生组织保留的细胞。
进一步研究表明,只有拟南芥莲座叶的侧芽才可能形成莲座叶,同样,茎生的侧芽只能形成茎生叶;通常拟南芥的萼片内不形成侧芽,但在AP1突变体中,萼片内侧基部出现了侧芽,且该侧芽只形成不同类型的花器官。
二、茎的分枝方式及其意义
植物的顶芽和侧芽存在着一定的生长相关性。
当顶芽活跃地生长,侧芽的生长则受到一定的抑制。
如果顶芽因某些原因而停止生长时,侧芽就会迅速生长,由于上述原因及植物的遗传特性,不同植物有不同的分枝方式(图6-4)。
(一)单轴分枝(monopodialbranching)(总状分枝)
单轴分枝是指从幼苗开始,主茎的顶芽活动始终占优势,形成一个直立的主轴,而侧枝则较不发达,其侧枝也以同样的方式形成次级分枝的分枝方式。
单轴分枝方式的植株呈塔形。
如红麻、黄麻等便是单轴分枝(图6-4)。
所以,栽培这类植物时要注意保顶芽,以提高其品质。
杨、松、杉等木本植物的树干高大挺直,是很有价值的木材。
(二)合轴分枝(sympodialbranching)(在栽培上,常采用整枝方法来调整果枝和营养枝(foliageshoot)的比例,在保证一定的营养面积的基础上,充分发挥果枝的增产作用,提高品质。
例如在棉花栽培中,当棉苗开始出现一、二个果枝时,把第一果枝以下的营养枝摘除,可以减少营养物质的消耗,促使果枝生长和花蕾发育。
当果枝上的花蕾达到一定数量时,把主茎和果枝的顶芽摘掉,留下一定数量的果枝和花蕾,以得到充分的营养供应,增加有效蕾数和铃重,有利稳产高产。
)
合轴分枝是指植株的顶芽活动到一定时间后死亡、或分化为花芽、或发生变态,而靠近顶芽的一个腋芽迅速发展为新枝,代替主茎生长一定时间后,其顶芽又同样被其下方的侧芽替代生长的分枝方式。
合轴分枝的主轴除了很短的主茎外,其余均为各级侧枝分段连接而成,因此,茎干弯曲、节间很短,而花芽较多。
合轴分枝在农作物和果树中普遍存在,如棉(图6-5)、番茄、马铃薯、柑橘类、葡萄、枣、李等的果枝(fruitshoot),茶树等在幼年期为单轴分枝,成长后则出现合轴分枝。
(三)假二叉分枝(falsedichotomousbranching)
假二叉分枝是指某些具有对生叶序的植物,如丁香、石竹、檞寄生等,其主茎和分枝的顶芽生长形成一段枝条后停止发育,由顶端下方对生的二个侧芽同时发育为新枝,且新枝的顶芽与侧芽生长规律与母枝一样,如此继续发育形成的分枝方式。
这样的分枝在外表上形似二叉分枝,实际它和顶端分生组织本身分裂为二所形成的真正的二叉分枝(如地钱)不同。
(四)分蘖(tiller)(禾本科植物的分蘖又可分为①疏蘖型:
分蘖节之间的距离较远,均在地下茎节上形成,各分蘖在地上的部分(分枝)呈疏松的丛生状,如小麦、水稻等;②密蘖型:
分蘖节是在靠近地面或地上部分形成,分蘖节间距短,呈紧密的丛生状,如狐茅属、针茅属等植物;③根茎型:
一些多年生具有根状茎的植物,如竹类、甘蔗、芦苇等。
地下茎上的侧芽开始生长时与主轴垂直,并以合轴分枝方式形成根状茎和不定根群,生长一段后,才向地上形成分蘖,如此类推。
)
分蘖是指植株的分枝主要集中于主茎的基部的一种分枝方式。
其特点是主茎基部的节较密集,节上生出许多不定根,分枝的长短和粗细相近,呈丛生状态。
典型的分蘖常见于禾本科作物,如水稻、小麦等的分枝方式(图6-6)。
第三节 茎的解剖结构
一、茎尖分区
茎的顶端叫做茎尖(stemtip),是由叶芽(顶芽)活动形成的。
顶芽活动时,生长锥的原分生组织分裂,向下产生初生分生组织,初生分生组织经初生生长形成初生结构,从而形成茎尖。
茎尖与根尖一样具有一定的形态结构特征,也可人为地划分为分生区、伸长区和成熟区三个部分(图6-7A)。
茎尖细胞不断进行分裂、生长和分化,使茎不断伸长并不断产生新的枝叶。
茎尖的生长分化过程与根尖基本相似,但由于茎尖所处的环境以及所担负的生理功能不同,其形态结构也有所不同:
茎尖没有类似根冠的结构,然而分生区的基部次第形成了一些叶原基(有时有腋芽原基)和幼叶,以及未发育成的节和节间,增加了茎尖结构的复杂性。
(一)分生区
茎尖分生区又称生长锥,一般为半球形,由一团具有分裂能力的原分生组织所构成。
茎尖顶端以下有叶原基和腋芽原基,有的还有芽鳞原基。
以原套一原体学说或组织细胞分区学说来说明茎尖生长锥的结构和分化动态。
1.原套-原体学说
原套-原体学说将茎生长锥分为原套(tunica)和原体(corpus)两部分。
原套位于其表面,由一层或数层排列整齐的细胞组成。
它们进行垂周分裂(anticlinaldivision),扩大其表面积。
原体是原套内侧的一团不规则排列的细胞,它们可沿着各种方向进行分裂,增大体积(图6-7A)。
在营养生长过程中,原套和原体的细胞分裂活动,互相配合,故茎尖顶端始终保持原套、原体的结构。
大多数的双子叶植物,原套通常是二层,而单子叶植物则有一层或二层。
观察表明:
当营养生长向生殖生长过渡时,茎尖的原套层数和原体的相对体积都会发生变化。
原套层数还可随季节、植株生长状况、发育阶段等的不同而改变。
2.细胞组织分区学说(不同植物的细胞组织学分区的具体特征不同。
茎尖顶端分生组织比较均匀的植物一般没有分区现象。
在茎尖顶端后部,周缘分生组织(周围区)和髓分生组织(髓区域)逐渐分化为三种初生分生组织。
在周缘分生组织内侧维管组织开始以原形成层的方式出现,其细胞因为纵向分裂占优势而变得比较细长,它们可与比较短而宽的基本分生组织的细胞相区别。
同时,原表皮也逐渐更多地显出成熟表皮的特征。
)
根据细胞学特征和组织分化动态的观察,在裸子植物和已研究的大多数被子植物中发现茎端有分区现象(图6-7)。
在原套、原体的中央部位,各有一个原始细胞群,前者称为顶端原始细胞区,后者的称为中央母细胞区。
这些细胞较大,并具较大的核和液泡,染色比周围的原套、原体细胞浅,细胞分裂较为频繁。
这两群原始细胞分裂形成了围绕在它们周围的周围分生组织区(peripherialmeristemzone)和下方的肋状分生组织区(ribmeristemzone)。
肋状分生组织区又称髓分生组织区(pithmeristemzone),其细胞较周围分生组织细胞更液泡化,主要进行有规律的分裂,因而形成特殊的“肋状”。
有些植物在肋状分生组织区和周围分生组织区上方还有整体如浅盘状的形成层状过渡区(cambium-liketransitionzone),其细胞在茎端纵切面上为扁平状,排列整齐,如同形成层及其衍生细胞所形成的形成层带,如菊等植物的粗壮茎端。
形成层状过渡区在叶原基形成的间隔期可以有体积的变化。
分生区下方形成初步分化的初生分生组织;由原套的表面细胞分化的原表皮层,周围分生组织和肋状分生组织分化形成的基本分生组织和原形成层(图6-8)。
(二)伸长区
茎的伸长区的细胞学特征与根相似,但该区常包含几个节与节间。
其长度可随环境改变。
二年生和多年生植物在进入休眠期时,伸长区逐渐变为成熟区。
伸长区由原表皮、基本分生组织、原形成层三种初生分生组织分化出一些初生组织,其细胞的有丝分裂活动逐渐减弱,伸长区可视为顶端分生组织发展为成熟组织的过渡区域。
(三)成熟区
成熟区的解剖特点是细胞的分裂和伸长生长都趋于停止,各种成熟组织的分化基本完成,具备了幼茎的初生结构。
在生长季节里茎尖的顶端分生组织细胞不断地进行分裂、伸长生长和分化成熟,结果使节数增加,节间伸长,同时产生新的叶原基和腋芽原基。
这种由于顶端分生组织的活动而引起的生长称为顶端生长(apicalgrowth)。
二、双子叶植物茎的解剖结构
(一)双子叶植物茎节间的初生结构
双子叶植物茎节间的初生结构可分为表皮、皮层和维管柱三部分(图6-9)。
1.表皮
表皮是幼茎最外的一层细胞,由初生分生组织的原表皮发育而来的初生保护组织。
表皮包括表皮细胞、气孔器、各种表皮毛。
表皮细胞形状较规则,排列紧密、相互嵌合,细胞一般不含叶绿体,有的含有花色素苷;细胞外壁厚、有角质层,有的还有蜡被,可起着保护和控制蒸腾,防止水分过度散失的作用。
有利于幼茎内的绿色组织进行光合作用。
如蓖麻(RicinuscommunisL.)、油菜(BrassicanapusL.)等。
表皮毛有单细胞和多细胞的,形状多样,具有加强保护的功能,有的表皮有腺毛或异细胞
 升级会员
升级会员 