人类的恐惧因何而来.docx
《人类的恐惧因何而来.docx》由会员分享,可在线阅读,更多相关《人类的恐惧因何而来.docx(15页珍藏版)》请在冰豆网上搜索。
人类的恐惧因何而来
人类的恐惧因何而来
要回答恐惧(fear)的来源,应该从三个关键的方面入手:
∙恐惧的PhylogeneticContinuity(种间一致性)
∙恐惧的基本分类
∙恐惧的神经基础
人类的情绪,特别是高级复杂的情绪,有许多仅仅是一种心理性/社会性的构造.意即,这些情绪是在社会/文化环境的影响下衍生而出的.然而恐惧不同.与恐惧密切相关的脑区:
amygdala(杏仁核),PAG(PeriaqueductalGrey)等,都位于新皮层之下,属于早期发展出来脑结构之一.恐惧因此是一种“上古情绪”.
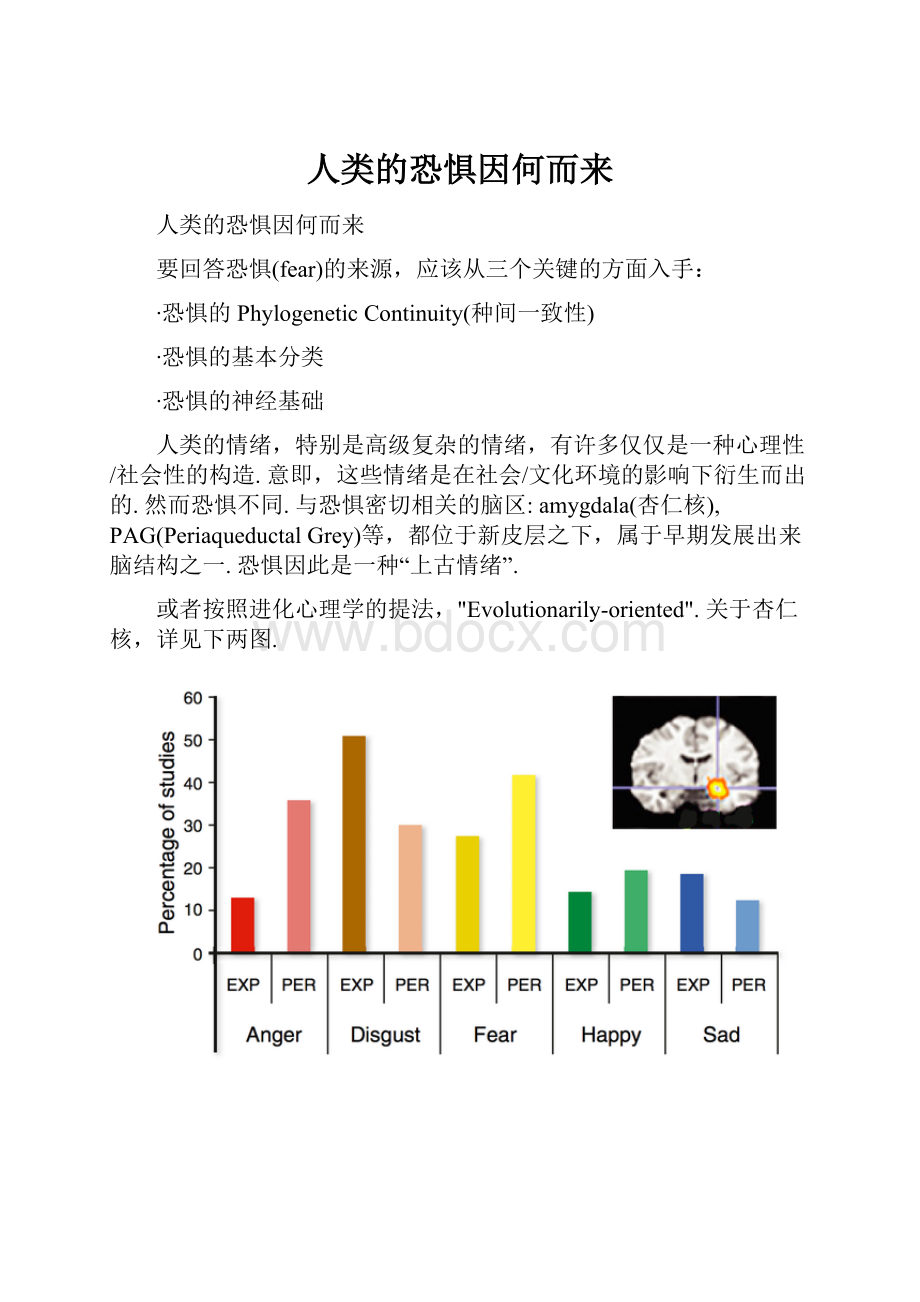
或者按照进化心理学的提法,"Evolutionarily-oriented".关于杏仁核,详见下两图.
Fig.1.2图片截取自[1].本图想说明的问题是,杏仁核绝不是恐惧的专属脑区.事实上,有关愤怒,恶心,悲伤,甚至愉悦相关的任务都在杏仁核有报告激活.
为什么会如此?
因为恐惧是为了生存.想象一下,如果我们的先祖遇到一头大野猪,不赶快跑路(fear-avoidance)或者挥舞棒槌将其锤死(fear/aggression-approach)的话,他就像破抹布一样为野猪的进化铺路了.事实上,包括恐惧,恶心(disgust),侵略性(aggression)在内的几种基本情绪,都是原初survivalcircuit(生存回路)的一部分.
进化心理学这谭水比较深,这里不赘述.感兴趣的同学可以参看[2]的第3,10章,以及[3].
恐惧的种间一致性
由于恐惧的作用如此重要,它在不同的物种中广泛存在.老人家达尔文是研究此话题的先驱.他在划时代的[4]中指出:
包括人类,非人灵长类,其他哺乳类,啮齿类,甚至无脊椎类在内的动物,对于恐惧,恶心,侵略性这几种情绪的表达,都有着功能上和行为上的高度相似性.这就是所谓的PhylogeneticContinuity(种间一致性).
我想表达的是动物与人类有相似的表达恐惧的行为.
这为什么重要呢?
因为只有这样,我们才能全方位无死角地研究恐惧的来源.心理学家和生态学家可以分别对人类/其他物种的行为进行研究,对恐惧进行明确的定义/分类;神经科学研究者则可以通过非侵入性的观测(人类)和侵入性的观测/解剖(动物)来考察恐惧的神经基础.两个方面相结合,我们就能得到关于恐惧的整个图景.Psychology,neurology,ecology,三位一体!
事实上,这也是恐惧相比其他人类复杂情绪,其研究成果更加深入,缜密的原因.
为了不把恐惧研究做成玄学,我们应该尽可能地剥离社会,文化与语言的影响,研究恐惧在物种间共通的本质,或者说,来源.
一点私货:
在心理学/认知神经科学研究可重复性问题Pantsonfire的情况下,适度的Reductionism(还原主义)是必要的.恐惧的具体表现形式诚然有很多,但他们都位于同一套“恐惧系统”之下.我们不能为每一种独特的恐惧都定义一套新的系统,甚至牵强附会地为它们附上不同的脑区.
RalphAdolphs在[1]中指出,要使包括恐惧在内的情绪研究真正成为一门科学,应该广泛地使用functionaldefinition(功能性定义).要做这个定义,第一步应该竖立的观念是:
恐惧(fear)!
=害怕(beingafraid)
我(也许)永远也不可能知道家里那只猫在被拉去洗澡的时候是不是“感觉到”害怕了.我也不知道那只苍蝇在被我拍死之前是不是“感觉到”害怕了.我更不可能知道让一只Toad在海上赛艇,它是否会“感觉到”害怕.对于人类来说,恐惧也不一定是与知觉相联系的:
如果恐惧刺激的出现十分迅速(比如B级片的jumpscare),人在意识到自己“害怕”之前就会有相应的生理反应.因此,仅从人类的“感觉”,或者说知觉层面描述恐惧是非常局限的.首先要明白,情绪并不等同于主观感受-情绪是大脑对于某些刺激的反馈,以及它引起的生理反应,以及最后所带来的人类行为的集合体.
“感到害怕”只是恐惧状态的产物/证据而已,并非恐惧本身."Evidenceforthestateisnotthestateitself"[1](状态的证据并不是状态本身).
弄清楚了这一点,我们就可以对恐惧进行功能性定义了.[1]中提出了一个操作性非常强的定义,"fearisaninterveningvariablebetweensetsofcontext-dependentstimuliandsuitsofbehavioralresponse"(情绪就是情境关联的刺激与相应的行为产出的中间变量).如下图所示.
4_r.jpg">Fig.3图片为自制.左侧是系统的输入,右侧是系统的输出.输入的是掠食者,疼痛,同类的侵略性等外界刺激.当然,这些刺激中也包括更加抽象,更加社会化的刺激,例如大家喜欢提到的“对未知的恐惧”,又或是F.D.R.的名言“
Fig.3图片为自制.左侧是系统的输入,右侧是系统的输出.输入的是掠食者,疼痛,同类的侵略性等外界刺激.当然,这些刺激中也包括更加抽象,更加社会化的刺激,例如大家喜欢提到的“对未知的恐惧”,又或是F.D.R.的名言“Onlythingwehavetofearisfearitself”.输出的则是各种行为,比较典型的有著名的FFF[5](别笑,这里是逃跑/战斗/凝滞).恐惧就是处在两组变量之间的黑箱子,是神经反应与生理变化的结合.
有了可以操作的功能性定义,我们可以整理一下思路了.应该如何着手对恐惧的研究?
恐惧的“来源”,归根结底是生物学意义上的(biologicalfoundation).对于实验科学而言,一个可能的思路是,想办法穷尽输入与输出的组合,研究这些组合中间的黑箱子(神经机制)到底长什么样子,并将它们归结到一个大一统的“恐惧的神经系统”的框架之内.
遗憾的是,人类毕竟比动物更“进化”.类别迥异的恐惧已经有了各自相对dedicated(专一)的神经回路,很难大一统到一个框架之下.还原主义不代表强行还原.在剔除了可以剔除的枝叶之后,我们慢慢啃那些难以撼动的分叉.
研究者们退而求其次,将恐惧按照不同的基本分类方法区分开来,并由此搞清楚了一些很有价值的与恐惧相关的神经机制.
恐惧的基本分类
在前文中,我们已经明确了恐惧的种间一致性,以及恐惧需要放在这个框架下研究的必要性.现在我假设大家接受了这种还原主义的立场,罗列学界几种比较基本的恐惧分类方法.
1.按刺激类别(获得方式)划分
刨去所有社会,文化因素的装饰,恐惧按照获得方式其实可以简单地分为两类:
与生俱来的(innate)与后天学习的(learned).
对于前一类,生态学家已经给出了答案:
feartowardspredator,feartowardsaggressiveconspecific(对掠食者,以及侵略性强的同类的恐惧).这一类的恐惧可以看作是hardwired(硬件的),或者说是在进化过程中印刻在基因中的.
而对于后一类,基本可以由巴甫洛夫那一套fearconditioning来解决(为无数的大鼠默哀一下).而最具有代表性的反射刺激就是疼痛刺激.
1)conspecificfighting(同类间的侵略性行为)2)典型的大鼠fearconditioning(恐惧反射)范式3)兔子对捕食者大猫的恐惧.
Fig.4图片来源于网络.从左上起,顺时针:
1)conspecificfighting(同类间的侵略性行为)2)典型的大鼠fearconditioning(恐惧反射)范式3)兔子对捕食者大猫的恐惧.
基于此,Gross和Canteras在[6]中将恐惧分成了
1)对掠食者
2)对侵略性强的同类以及
3)对疼痛刺激
的恐惧,并分别挖出了对应的神经回路.详见下一节.
2.按时间属性划分
按照时间方面的属性状态(其实这么说很不准确,但我似乎找不到更好的中文表达法:
/),恐惧(fear)其实可以分为恐慌(panic)和焦虑(anxiety).前者是短暂的(phasic),激烈的,由某个显性的外界刺激引起的状态;后者则是持续的(sustained),隐式的,跟准备与预期相关的状态.
这种区分很好理解.我们可以假设解一道数学题的情况.假如你正在考场上,面对着一道解不出来的数学题.这个时候你的情绪归类于恐慌.因为题就在你面前,且解不出题的状况正在当下;另一方面,如果你在考前前一晚,惦记着一道老师说过会考,而你还不会的数学题,这个时候你的情绪就归类于焦虑.因为题是隐性的,解不出题的情况只是你的预期.
Davis等人在[7]中总结了依照这种分类方法在人类与大鼠中的研究.神经回路详见下一节.
3.按空间属性划分
恐惧在空间上的分类最初是由生态学家们提出来的.这个空间属性是针对被掠食者与掠食者的位置关系而言的.Fanselow和Lester[7]提出了如下图所示的理论模型(低清瞎眼注意:
whatever),主要成就在于清晰地定义了掠食者从远道近接近被掠食者的各个过程以及可能对应的行为.自上而下分别是:
Nopredatorpotential(完全安全),Pre-encounterphase(掠食者已经在周遭环境的某处出现,但是还未进入追击状态),Post-encounterphase(掠食者开始追击被掠食者),Circa-strikephase(掠食者正面攻击被掠食者的瞬间).
Fig.5(这文章太老了,实在没有更清晰的图了)这个模型被称为Predatoryimminencecontinuum(掠食迫近连续统?
whatever),主要成就在于清晰地定义了掠食者从远道近接近被掠食者的各个过程以及可能对应的行为.自上而下分别是:
Nopredatorpotential(完全安全),Pre-encounterphase(掠食者已经在周遭环境的某处出现,但是还未进入追击状态),Post-encounterphase(掠食者开始追击被掠食者),Circa-strikephase(掠食者正面攻击被掠食者的瞬间).
虽然这只是一个生态学的理论概念,并且大多运用于该领域predator-preyinteraction(掠食-被掠食者互动)的研究(生态学的这个话题其实很有趣,改天另开一坑),但Mobbs等人将此概念运用到人类脑成像研究中,取得了一些洞见.相关神经回路依然见下一节.
3.其他分类方式
LeDoux将恐惧这个情绪放到一个统合的"Survivalnetwork"里面研究,因为个人觉得其理论模型比较臃肿(没说他不好,轻拍),这里不赘述.感兴趣的读者请参看[9].
恐惧的神经基础
我们终于来到了最后一步的重头戏:
根据我们划好的功能性恐惧分布,来探究恐惧的神经基础:
当名为恐惧的情绪发生时,我们大脑里到底在干什么?
1.刺激类别划分的神经机制
Gross和Canteras在[6]中总结出:
对掠食者的恐惧(innate),对同类侵略性行为的恐惧(innate),以及对疼痛刺激进行反射学习的恐惧(learned)分别对应这三条神经通路:
从上自下分别为
Fig.6引自[6].从上自下分别为
对掠食者恐惧的通路=LA(lateralamygdala)--->pBMA(posteriorbasomedialamygdala)--->pvMEA(posteroventralmedialamygdala)--->dmVMH(dorsomedialVMH)--->vlPMD(ventrolateralPMD)--->dlPAG(dorsolateralPAG).
对同类侵略行为恐惧的通路,对疼痛刺激进行反射学习的通路:
与上面的通路有相似之处,由于一般读者不会对这些细节感兴趣,这里不再赘述,细节见[6].
可以清楚地看到,除了杏仁核(amygdala)之外,海马(hippocampus),隔膜(septum),下丘脑(Hypothalamus)以及PAG(好吧...中脑导水管周围灰质)等区域也参与到了恐惧的处理之中.这些区域之间也有着较为复杂的交互机制.
记得最开始看到这个review的时候,我是挺开心的.这个的takehomemessage在于:
结合行为/电生理/解剖研究,我们可以清楚地定位出几种基本恐惧类型分别的神经通路,哪怕它们的区别十分微小.
2.时间属性划分的神经机制
Davis等人在[7]这篇雄文中,结合人类与大鼠的相关研究,提出了一个关于CeA(CentralnucleusofAmygdala,杏仁核中间部分的核...)以及BNST(bednucleusofthestriaterminalis,终纹床核)的工作模型,用来解释时间尺度上不同的恐惧:
恐慌(panic)和焦虑(anxiety)是如何产生的.
Fig.7引自[7]按照此工作模型,恐惧相关的刺激首先快速激活BLA(basolateralamygdala)与medialCeA,引发快速的恐慌反应(panic).另一方面,向lateralCeA的输入会向BNST释放CRF(促肾上腺皮质激素释放因子),引发一个更加缓慢,持久的焦虑反应(anxiety).来自BNST的抑制性反馈作用也可以终止恐慌反应,从而完成自恐慌向焦虑的平滑切换.
这在一直傻傻分不清楚的恐慌和焦虑之间切了一刀,我认为是很有意义的工作.
3.空间属性划分的神经机制
Mobbs将生态学中掠食者威胁连续统的概念嫁接到了人类fMRI研究之中.他认为当威胁较为抽象或者空间上远离时,可以对应Post-encounterphase(掠食者已出现,但刚刚开始追逐),而当威胁极近时,可以对应Circa-strikephase(掠食者攻击得瞬间).因此他设计了一系列实验来研究人类被试处于这两种状态,以及在两种状态间转换时的大脑工作方式.
在[10]中,为了达到恐惧刺激的效果,实验者让人类被试以为自己的脚边不远放了狼蛛(tarantula).事实上是事先录好的录像带啦.实验装置如下图所示.
Fig.8截取自[10].当被试躺在MRI扫描仪中时,实验者给被试播放事先录好的录像带,被试会看到狼蛛被放进了1-5中的某一个格子中,或者是从其中一个格子被移动到另外一个格子.这样一来,就制造了危险从空间上接近/远离的效果.(其实如果能战胜IRB,好希望直接放真正的蜘蛛进去wink)
Fig.9截取自[10].A:
当狼蛛慢慢接近被试时,包括dACC(dorsalanteriorcingulatecortex,背侧前扣带皮层)和Midbrain(中脑,含PAG)在内的脑区参与了这一过程.B:
另一方面,当狼蛛慢慢远离被试时,包括PCC(posteriorcingulatecortex,后扣带皮层)和omPFC(中前额叶皮层)在内的脑区参与了这一过程.
实际上在这之前,Mobbs等人已经用电脑游戏范式得出了类似的结论(参看[11]).这些研究在一定程度上深化了我们对于恐惧在空间属性上处理的理解.
结语
限于篇幅(以及精力...困了准备躺尸)所限,这里不打算将恐惧这一话题全部展开.但对于“恐惧的来源"这一问题,本文已经从操作性定义以及生物基础上给出了一定的解答.
我们离完全理解恐惧还比较远-特别是缺乏computational的研究.当然,我也不希望早早地把这个问题搞清楚,那样我就没办法对被试做过分的事情了:
)
P.S.应该会扩写成文章
-----------------------------------一般分割线---------------------------------
@梁正源问:
为什么刚出生的婴儿也会知道恐惧啊?
抱着的时候失手往后一靠,他一脸的惊恐表情,这么小啥都不会都会怕摔
答:
文中已经提到了,因为恐惧是人类为了生存,保有的最基本的情绪(可以说没有之一),属于genetically-hardwired,因此刚出生的小婴儿也会对失重/剧烈的高度差变化感到恐惧.您抱他的时候失手,他会害怕,并不奇怪.
不过小婴儿的对危险刺激的反应其实是很弱的(与成人相比).从行为上讲,这是因为他们对于危险刺激的种类还不甚了解.从神经机制上讲,一方面是杏仁核本身还处于发展之中,另一方面也因为前额叶-杏仁核的调控连接还未完全形成.理论上推测,这是一种进化过程中形成的“探索试错机制”(exploringmechanism):
因为婴儿幼小的时候有双亲保护,让他们在对危险刺激不太敏感的情况下尽情探索环境,对未来的发展是有益的.有发展心理学的研究证实了前额叶-杏仁核的静息功能连接随着年龄增加而减弱,而这种静息态连接的减弱正反映了任务态连接的逐渐增强,从一定程度上证明了上述理论.前额叶对杏仁核的调控正是在外界不断输入的恐惧刺激中慢慢增强的.具体可以参看Gabard-Durnametal.2016(J.Neurosci.).
[1]Adolphs,R.(2013).Thebiologyoffear.CurrentBiology,23
(2),R79-R93.
[2]Buss,D.(2015).Evolutionarypsychology:
Thenewscienceofthemind.PsychologyPress.
[3]LeDoux,J.E.(2012).Evolutionofhumanemotion:
aviewthroughfear.Progressinbrainresearch,195,431.
[4]Darwin,C.(1872).1965.Theexpressionoftheemotionsinmanandanimals.London,UK:
JohnMarry.
[5]Bracha,H.S.(2004).Freeze,flight,fight,fright,faint:
adaptationistperspectivesontheacutestressresponsespectrum.CNSspectrums,9(09),679-685.
[6]Gross,C.T.,&Canteras,N.S.(2012).Themanypathstofear.NatureReviewsNeuroscience,13(9),651-658.
[7]Davis,M.,Walker,D.L.,Miles,L.,&Grillon,C.(2010).Phasicvssustainedfearinratsandhumans:
roleoftheextendedamygdalainfearvsanxiety.Neuropsychopharmacology,35
(1),105-135.
[8]Fanselow,M.S.,&Lester,L.S.(1988).Afunctionalbehavioristicapproachtoaversivelymotivatedbehavior:
Predatoryimminenceasadeterminantofthetopographyofdefensivebehavior.
[9]LeDoux,J.(2012).Rethinkingtheemotionalbrain.Neuron,73(4),653-676.
[10]Mobbs,D.,Yu,R.,Rowe,J.B.,Eich,H.,FeldmanHall,O.,&Dalgleish,T.(2010).Neuralactivityassociat
 升级会员
升级会员 