土壤植物大气连续体.docx
《土壤植物大气连续体.docx》由会员分享,可在线阅读,更多相关《土壤植物大气连续体.docx(35页珍藏版)》请在冰豆网上搜索。
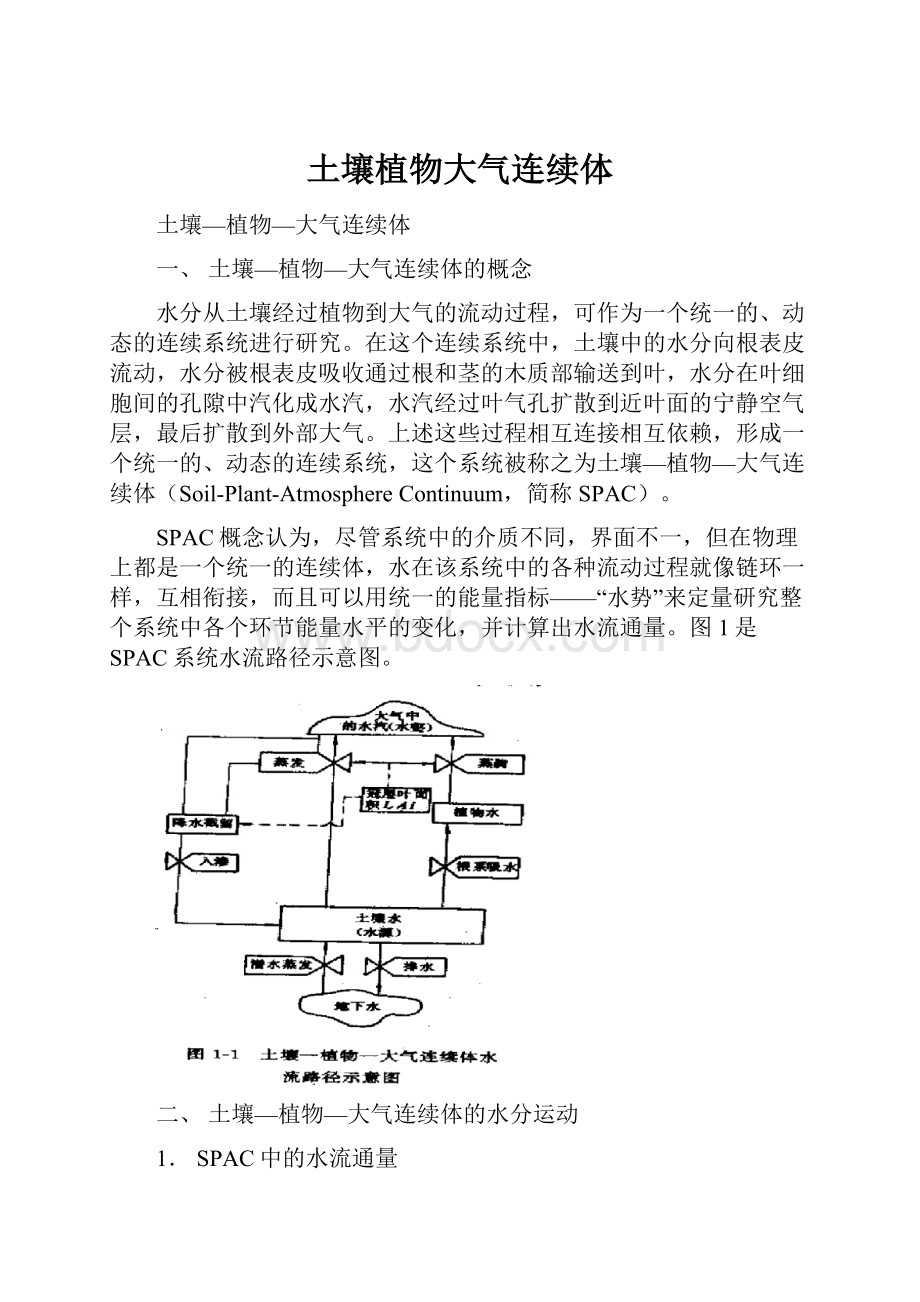
土壤植物大气连续体
土壤—植物—大气连续体
一、土壤—植物—大气连续体的概念
水分从土壤经过植物到大气的流动过程,可作为一个统一的、动态的连续系统进行研究。
在这个连续系统中,土壤中的水分向根表皮流动,水分被根表皮吸收通过根和茎的木质部输送到叶,水分在叶细胞间的孔隙中汽化成水汽,水汽经过叶气孔扩散到近叶面的宁静空气层,最后扩散到外部大气。
上述这些过程相互连接相互依赖,形成一个统一的、动态的连续系统,这个系统被称之为土壤—植物—大气连续体(Soil-Plant-AtmosphereContinuum,简称SPAC)。
SPAC概念认为,尽管系统中的介质不同,界面不一,但在物理上都是一个统一的连续体,水在该系统中的各种流动过程就像链环一样,互相衔接,而且可以用统一的能量指标——“水势”来定量研究整个系统中各个环节能量水平的变化,并计算出水流通量。
图1是SPAC系统水流路径示意图。
二、土壤—植物—大气连续体的水分运动
1.SPAC中的水流通量
在SPAC中,水分运动的驱动力是水势梯度,即水总是从水势高处向水势低处流动。
系统中水流动的速率与水势梯度成正比,与水流的阻力成反比。
由于在SPAC中各个部位的水流阻力和水势并非是恒定不变的,严格地说SPAC中的水流是非恒定流。
但在实际中,一般忽略植株体内储水量的微小变化,认为SPAC中的水流为连续的恒定流。
按照Huber(1924),Gradmann(1928)和vandenHonert(1948)的假说,SPAC被描述成类似欧姆系统中的电流。
因此,任何一段液流中的流量为:
(1)
式中:
q为流量,以质量/时间为单位表示;-Δψ为势能降,以能量单位表示;R为阻力。
vandenHonert(1948)在描述由于水势梯度而造成水分运动的链式理论时,将系统表示为:
(2)
式中:
ψs、ψr、ψx、ψl、ψa—分别为土水势、根—土壤界面的水势、根木质部内的水势、叶水势和大气水势。
Rs、Rr、Rx、Ra—分别为水分流经土壤到根-土界面、根-土界面到根木质部、再从根木质部到达叶蒸发表面、最后从叶蒸发表面进入开放大气中的各阻力。
在一维稳定流条件下,可用图所示的电阻网络来模拟SPAC中的水流状况。
图2
中用包括土壤阻力、根阻力、茎阻力、叶阻力和空气阻力的串联电路代表图2(b)中复杂的水流路径。
蒸腾着的植株根部四周土壤的水势通常在0—-1.0MPa的范围内,叶水势通常为-0.2—-2.0MPa,作物在严重水分胁迫下可达-3.0MPa,某些耐旱植物可能低至-5.0MPa。
若用方程ψa=RTkln(e/es)(R为气体常数,Tk为绝对温度,e/es为相对湿度)把空气相对湿度换算成相等的水势,则相对湿度在98%一48%范围内,相当于大气水势约为-10~-100MPa,在干旱半干旱地区空气相对湿度往往低于48%,因此其大气水势还会低于-100MPa。
由此可知,从土壤到叶片的水势降低约为1.0MPa或更少,但从叶到大气则降低几十甚至达100MPa。
这表明,在植株的水分吸收和蒸腾这样一个耦合过程中的主要阻力是从叶到大气的阻力,即蒸汽相的阻力最大。
经过许多人发展的VandenHonert假说具有普遍意义。
由于主要的、控制性的阻力在叶一气系统之间,这意味着,通过整个连续体的通量,主要决定于调节蒸腾的那些因素,包括供应水分汽化的能量。
然而,值得注意的是,事实上土壤水分枯竭会显著地降低蒸腾,上述结论的明显矛盾,是由于吸收途径上的反馈条件改变了蒸腾途径上的阻力。
根部通常对水分流动有相当大的阻力,大部分与从表皮经过皮层到中柱的通道有关系,在皮层中由于在内层细胞中的凯氏带的作用,它处于一定程度的生理控制之下。
为了使吸收水分的速率与植物蒸腾速率近似相等,在根系传导系统中所需的压力差大约从0.44MPa到5.0MPa不等(Kramer,1969)。
一般认为,根部阻力是导致蒸腾与根系吸水之间时间滞后现象的原因,这种滞后经常在迅速蒸腾的时期中引起叶片水分不足和气孔开度减小或关闭的现象。
由上可知,虽然vandenHonert的这种处理方法提供了一个模拟SPAC中水流的实用模型,但它确实过分简化了。
首先,它假定SPAC中的水流是稳定流,但在植物体中是很少存在的,因此,严格地说它是不适用于测定时间短于在植株体内形成稳态流所需要的时间的情况;其次,它假定在运输途径的各部分(例如在根中)的阻力是恒定的,但试验表明根中阻力有时是随着流速而改变的;第三,水以液态通过土壤和植株运行,但在叶气孔腔内转变成水汽,虽然运移的驱动力始终是化学势差,但液态流直接与水势差成比例,而水汽扩散则与水汽压或水汽的浓度差成比例,由水汽压把湿度梯度转换成相当的水势梯度,则大大地夸大了水汽相中势能的下降,估算出的叶一气系统水流阻力的量级与实际有较大的差异.因此,有必要发展和改进vandenHonett的稳定流模式的假定,引入水容的概念,提出与植物水分状况相符的非稳定流模式。
象电容器一样,水贮存在薄壁组织细胞中用于补充蒸腾丧失的水分。
在叶片和茎的薄壁组织细胞中贮存的水量很大。
植物在炎热的晴天有时会失去它的原有含水量的25%一30%,但在晚上就能得到恢复。
在中等蒸腾的日子里,叶片含水量的日变化一般约为10%。
正是由于这种水容因素的存在限制了欧姆定律模拟的严格应用。
2.SPAC中根内的水分流动的推动力
vandenHonert(1948)采用欧姆定律类推植物体内的水流,把根内的水流途径区分为两部分,从表皮到木质部的径向途径和沿着木质部流动的轴向途径。
对于一个稳恒的液流来说,往往还加进土壤中的径向途径。
此时:
(3)
式中:
ψsm、ψrm、ψr、ψx、ψxh、ψbh—分别为土壤基质势、根—土壤界面的基质势;
ψr、ψx—分别为根—土壤界面的水势、根木质部内的水势,水势ψ=ψm+ψπ,ψm为基质势,ψπ为渗透势;
ψxh、ψbh—分别为根木质部内和根基部的水势(hydraulicpotential);水势ψh=ψp+ψg,为ψp压力势和为ψg重力势。
Rs、Rβ、Rα分别为单位土壤表面和根的径向途径β与根的轴向途径α的水流阻力。
上式说明了液流途径的性质不同将有不同的ψ组分在起作用。
由于土壤内溶质流动没有半透性屏障加以阻挡,所以ψπ的任何梯度都是无效的,而流向根的液态水仅仅是靠基质势(ψm)差推动的。
同样地,在木质部内轴向液流途径中也没有半透性屏障加以阻挡,所以木质部内水分与溶质的移动是在由压力势和重力势组成的水势梯度下进行的。
在一条垂直长度为L的根内,其流量f可以用下式表示:
(4)
式中:
Kα为轴向传导率。
其倒数为轴向液流阻力Rα=1/Kα。
在表皮与木质部之间存在着一个半透性屏障,这使得根的行为变的如同一个渗透计一样,从表皮到木质部导管的径向途径内的水流受两者之间由基质势与渗透势组成的水势所推动。
从表皮到木质部导管的径向途径内的水流量Jw可以用下式表示:
(5)
式中:
Kβ为径向传导率。
其倒数为径向液流阻力Rβ=1/Kβ。
3.SPAC中的水流阻力分布规律
水分通过SPAC遇到土一根系统、植物体和叶一气系统的阻力(如图3),主要包括土壤阻力、根系阻力、茎内和枝条的木质部阻力、叶肉阻力和叶片气孔、角质层阻力以及叶周围
宁静空气层和外界空气层的阻力。
1)SPAC中各部分水流阻力的计算
(1)土壤阻力
Gardner(1960)通过对单根稳态流的研究给出的土壤阻力Rs计算式,即:
(6)
式中:
K(θ)为土壤导水率;zr为根系深度;Lv为根长密度,即单位体积土壤中的根系长度;r为根半径。
该公式计算的土壤阻力单位用S表示。
该式需要根系密度资料,应用不方便。
Gardner和Cowan还提出了仅需土壤基质势计算土壤阻力Rs的另一个经验公式:
(7)
式中:
ψm为土壤基质势;ψm0为土壤水分特征曲线上饱和点的进气值,为-0.8m水柱。
用该式计算的Rs单位是s/m。
(2)根系阻力
土一根系统的总阻力Rsr包括土壤阻力Rs、根系吸收阻力(径向阻力)Rβ和根内木质部传导阻力(轴向阻力)Rα。
土一根系统的总阻力Rsr可由势差(ψsm一ψbh)与蒸腾速率T求得:
(8)
根系吸收阻力(径向阻力)Rβ能采用Herkelrath(1977)公式确定:
(9)
式中:
θs为土壤饱和含水量;θ实际含水量;rr为单位根长的水流阻力(s/m);Lz为单位面积土壤上的根长度(cm/cm2)。
该式计算的的单位是S。
根内木质部传导阻力R。
可根据圆形毛细管束中水流的Poiseuille方程导出计算公式:
(10)
式中:
ν为水的运动粘滞性系数,与温度有关,当温度为20℃时,ν=0.01007cm2/s;z为原点在地面、垂直向下的深度;N为单位面积上的根条数(cm-2);rH为深度内所有根系的有效根半径。
Ponsana发现在任何特定深度处根管的综合半径为:
(11)
式中:
n为该深度处取得根样数;ri为第i条根的半径。
一定深度范围内有效根半径可以按调和平均值来计算:
(12)
式中:
m为土壤分层数。
例如,在0到90cm深度区间内将根系分为三个土壤层次,其有效根半径为:
(13)
(3)植物体的传导阻力Rp
植物体的传导阻力包括植物根系阻力、茎和叶肉阻力,可表示为:
(15)
地上部分植物内部的传导阻力Rup则为:
(16)
(4)叶一气系统的水流阻力叶一气系统的水流阻力分布如图4所示。
角质层阻力rc相对于气孔阻力rst来说要大得多,叶片水分散失的主要路径是气孔。
叶肉阻力较小,可以忽略不计。
在叶气孔水分散失路径中的水流阻力主要是气孔阻力和叶片层流边界层及冠层上方湍流边界层的阻力。
叶一气系统的总水流阻力Rla可根据叶一气系统水势差和蒸腾速率求得:
(17)
需要注意的是以上各部分阻力的计算均是以vandenHonert的恒定流假定为基础的,Hailey等人(1973)、Bunce(1978)、Jones(1978)、Katerji和Hallaire(1984)及Passioura和Munns(1984)曾指出这种假定不总是能应用的。
例如,Bunce(1978)发现象大豆和棉花这样一些种类的作物蒸腾连续增加,而叶水势则保持不变。
各种各样的理论已被提出来解释这种现象。
Bunce认为由于根系延伸而引起的根系阻力减小可能减小植物体传导阻力值。
但是,Katerji和Hallaire(1984)则认为大的蒸腾需求使得植物释放其内部的贮水量,植物体阻力相对保持不变。
这种解释还得到B.H.1ynn和T.N,Carlson(1990)的赞同,因此,能够假定根一茎阻力在叶水势的某一变化范围内保持不变。
2)SPAC中各部分水流阻力的相对重要性
(1)植株体内各部分水流阻力的相对重要性
Jensen等(1961)通过测定向日葵和番茄各部分的阻力,得出向日葵根的阻力为叶片的1.6倍,而近于茎的4倍。
番茄的阻力较向日葵要低,但根的阻力为叶片的1.4倍,茎的4倍。
Neumann等(1974)根据不同蒸腾速率下,用在茎杆的不同高度测定的叶水势来估算水分运行阻力,计算出玉米茎、叶、根的阻力分别为植株总阻力的15%~25%,15%~30%和50%~70%。
但在向日葵中其茎、叶、根的阻力分别占总阻力的8%、20%和70%。
因此,在植物体中的水流阻力主要分布在根系部分。
(2)土壤阻力、植株体阻力和叶一气系统阻力的相对重要性
康绍忠根据在西北农业大学灌溉试验站测得的不同深度的土壤基质势、不同时间的植物
叶片水势、大气水势和蒸腾速率日变化资料计算了土壤阻力、植株体阻力及叶一气系统阻力。
表明,土壤基质势越低,土壤阻力越高,当土壤饱和或接近饱和时,其水流阻力可忽略不计。
冬小麦和玉米植株体的水流阻力在相近的生育阶段无明显的差异。
对于同一作物,如冬小麦在后期的Rp大于前期的2—4倍。
在作物相同阶段缺水处理和充分供水处理的植株体水流阻力相差不明显,但缺水处理的水流阻力略低于充分供水处理的,这可能是由于充分供水处理的根系主要分布于表层,而缺水处理的根系要向更深层次伸展所引起的,但这种差异要比Rs和Rla随土壤水分的变化小得多。
植株体阻力具有明显的日变化,每日上午时较低,夜晚要比中午的大2倍以上。
干处理的Rla明显大于湿处理的Rla,这主要是由于干处理的气孔阻力相对较大而引起的。
Rla具有明显的日变化,早、晚较大,中午较低;生育阶段后期的Rla大于前期的Rla。
SPAC中的水流阻力在叶一气系统最大,该部分的水流阻力占整个SPAC中的总水流阻力的97%以上,叶一气系统的水流阻力是植株体内传输阻力的50倍左右,是土壤阻力的几千倍。
因此,在SPAC中叶一气系统的水流阻力对控制系统中的水流运动起主导作用。
由于从土壤到叶气孔腔的水流是液态流,而从气孔扩散到大气的水流过程是汽态流,即系统中的水流过程产生了相变,因而对于在汽态流和液态流的阻力之间进行比较的合理性还值得研究。
由水汽压把湿度梯度转换为相当的水势梯度,大大地夸大了水汽相中势能的下降。
例如:
-3.0MPa的叶水势相当于30℃时叶片的相对湿度为98%或水汽压为41.5hPa。
空气相对湿度从98%降低到50%,水汽压约降低50%,但相当量的水势梯度则降低近25倍。
所以按照vandenHonert对欧姆定律的应用,大大夸大了叶一气系统中的水流阻力。
但由于在单位上的不同,当然也不能把以前从浓度梯度计算出来的水汽扩散阻力与从水势梯度计算出来的水汽扩散阻力等量齐观,因在叶片一空气路径中的水汽流与在木质部传导系统中的液态流之间没有什么明显的物理相似性。
所有的流动基本上是由于水势差而推动的,因此,用SPAC中各个部分的水势差推导其水流阻力的量级主要是为了实用的方便,在其理论上仍存在局限性。
三、SPAC系统中的水热动态
自从Philip1996年提出SPAC水流运动的概念后,SPAC水流运动的摸拟研究工作得到了很快的发展和不断的完善,许多人在这方面作了大量研究。
特别是象vandenHonert(1948)、Philip(1957),Monteith(1965),Cowan(1965)等人的研究工作为SPAC水流模型的发展奠定了基础。
Lambert和PenningdeVries(1973)提出的TROIKA模型,考虑了土壤水分的单根径向移动、单叶片的水汽和热量传输及根系吸水的作物阻力和土壤阻力;Lemon等(1973)提出的SPAC模型考虑了冠层内复杂的水汽、动量和热量廓线方程,Nimah和Hanks(1973)的SPAC模型考虑非稳定、非线性宏观源汇项等。
70年代后期有一些较大的模型出现,如Feddes等(1974)对Nimah和Hanks(1973)模型进行了改进,蒸散用综合方程,土壤蒸发模型通过辐射计算,根系吸水使用了根密度随深度的指数分布函数,Hasen(1975)提出并于1979年改进的称之为HEJMDAL模型的水流动态模型考虑了光合作用、呼吸和作物生长,构成了一个复杂的SPAC水流模型。
该模型的输入量有:
(1)土壤物理参数、土壤水分特征曲线、土壤导水率;
(2)作物参数:
根系生长分布、叶面积指数、作物水流阻力、作物生长(光合作用+呼吸作用);(3)气象参数:
总辐射、净辐射、温度、空气湿度、风速、降水量。
最重要的输出结果是:
蒸散、土壤和作物的水分状态和作物生长状况。
后来又相继出现子一些修改模型,如:
Federer(1979)考虑了土壤水势和根系分布而建立的估算根系水分运输阻力的理论模型,以及Camillo和Gurney(1983)、Bristow(1986)等提出的模型,这些模型无疑在涉及机制的前提下使其有—定的普适性,但大多数模型假设条件太多,缺乏详细田间实验验证,而且需要输入较多的参数,这是目前难于克服的一个困难,一些比较实用的模型又过于简化,与实际情况相差太远,不便于应用。
把点上的研究结果应用于面上是一个非常困难的工作,但却是十分必要的,因此就需要建立—个理论清晰严谨、实验基础扎实,即能充分反映SPAC水流各个环节的机理和过程又能使用常规观测资料进行预测预报的模型。
1.根区土壤水热运动模型
自从1856年法国工程师Darcy由实验得出水流运动的数学模型以来,土壤水流运动的数学模拟与实验研究工作得到不断发展,在此期间Darcy(1856),Richards(1931,1936)和Philip(1957,1958)等人的工作给土壤非饱和水运动模型的形成与发展奠定了基础。
1856年法国工程师Darcy通过实验得到被称之为达西定律的饱和土壤水流运动方程,开创了水流运动的定量研究。
为探索非饱和土壤水流运动的机理,布利格斯(1877年)提出了毛管假说},他将土壤孔道看成是一些大小不同的毛细管,认为表面张力是土壤水分保持的主要原因,1907年Buckinggham在此基础上,以物理学和热力学理论为基础,进而对毛管理论作了全面分析,首次提出了毛管势(即基质势)的概念,并依据欧姆定律得出了恒温条件非饱和土壤水流运动方程,为非饱和土壤水流运动研究打下了基础。
1936年Richards在前人工作的基础上,得出了被称之为Darcy和Richards非饱和土壤水流运动方程,该模型的提出标志着土壤水流运动模型研究进入了一个高级水平。
Darcy和Richards方程只是对恒温和均质土壤而言的,因此对温度变化较大和非均质性明显的土壤是不适用的。
实验指出对于细质土壤存在一种阀值水势梯度,低于这个梯度就没有水流发生(Swartzendruber,1962;Miller等,1963;Bolt等,1969),在这种情况下,Darcy和Richards方程就不能应用。
另外Darcy和Richards方程忽略了土壤中汽态水的运动,而非饱和带中汽态水的运动在土壤比较干的时候占水流量的很大一部分,且随着土壤含水量的减少,所占比例逐渐增大,因此在这种情况下不考虑非饱和带中汽态水的运动,将会给总水流量的计算结果带来明显的误差。
1957年,Philip在他的一系列出色工作中,结合前人的理论和实验考虑了非饱和带中汽态水的运动,首次得出了非恒温条件下的土壤液态和汽态水流运动方程,成为土壤水流模型发展的又一个里程碑。
Philip方程考虑了非饱和带中汽态水的运动,使水流运动方程前进了一大步。
但Philip
方程仍未考虑土壤的滞后效应,因此只适合单一的土壤水分变化过程。
另外,在非饱和带中,另一个与土壤水流运动有关的是有结构土壤,即土壤中所包含的一些较大的,多少相互联系的孔隙。
在这种土壤中,土壤的水流运动发生较大变化。
Philip方程也没有考虑土壤的空间变异和有结构性,所以在这种情况下,也是不适用的。
deVire于1958年对Philip方程进行了扩充,引进了吸附水中的热平流和湿润热。
随后Jackson(1972)、Geoenevelt等(1974)、Roots(1975)、Camillo(1986)等人进一步作了补充。
1979年Sophocleous将Philip方程改为以基质势为变量的方程,在此基础上Milly(1982)考虑滞后效应得出了非均质土壤水热联合运动模型。
关于土壤的空间变异,虽然近期已在这方面作了大量研究,但目前仍未有比较理想的模型。
在一般情况下水平方向水流运动与垂直方向水流运动相比可忽略不计,因此在田间研究时一般多采用一维水流运动方程。
由Philip和deVire(1957)提出并经后来—些学者(Jackson等,1974;Hammel等,1981;
Camillo等(1983)多次修正的一维水热运动方程为:
(18)
(19)
上式中qW、qH分别为土壤中的水分和热量通量,Ts为土壤温度,ψ为土壤水基质势,KWψ、
KWT分别为取决于水势和温度的土壤非饱和导水率,KHT、KHψ分别为取决于土壤温度和水势的土壤导热率。
据Camillo等(1983)和Mclnnes等(1986):
(20)
(21)
(22)
(23)
式中KW为土壤中液态水的传导率,KWTV为土壤中汽态水的传导率,DVa为水汽在大气中的扩散系数,fa为土壤有效孔隙度,ρ0为饱和水流密度,η是放大系数(据Mclnnes等,1986为1.021),g是重力加速度,h为相对湿度,R是汽体常数,据Mclnnes等,1986:
(24)
(25)
(26)
(27)
式中ε是土壤孔隙度,α是小于1的常数(2/3),据Kimpball(1986):
D0=0.229cm/s,R0=6.0035,R1=4975.90K
据守衡方程,当没有源汇项时,各层土壤的平衡方程为:
(28)
(29)
式中CW、CH分别为土壤中的比水容量和比热容量。
结合(6)式和(7)式有:
(30)
(31)
对于从土壤到作物到大气的水分运动,根系吸水是重要环节,当有根系时,各层土壤的平衡方程为:
(32)
(33)
式中SW是单位土体中根系的吸水量,这就是有作物情况下根区土壤的基本水热运动方程,其中的根系吸水项SW是连接SPAC水流运动的关键环节。
由于根系吸水的确定非常困难,因此目前国内的大多数计算模型和国外的一些计算模型都不考虑根系的吸水项,而总是把它与蒸发一起作为边界条件来考虑,这样不但模拟结果与真实情况相去甚远,而且也不能完全揭示作物体内和整个SPAC中的水流运动过程,因此也不能真正了解作物的需水本质并对农田水分进行调控。
模型应全面考虑SPAC水流运动的各个环节和过程,力求建立比较完整的能比较准确反映实际情况的模型。
2.作物蒸散模型
蒸腾是作物体内的水分通过气孔向大气散发的过程,与作物的各种水分过程系密切相联。
精确地估算蒸腾量是SPAC水流环节中难度很大的课题。
1953年Penman首次提出了计算单个叶片气孔蒸腾的模式,1959年Couvey把气孔阻力的概念推广到整个植被表面,1965年Monteith在Penman和Couvey等人的工作基础上得出了计算整个冠层、被称之为Penman-Monteith公式的模式。
该模式全面的考虑了影响作物蒸腾的大气因素和作物生理因素,为蒸腾和蒸散的研究开辟了一条新途径。
蒸腾包括两个阶段,液态水在气孔腔内汽化,汽化了的水分子通过气孔和大气边界层进入天气。
液态水汽化需要能量,对于给定的蒸腾面,其能量平衡并程为:
(34)
式中RN为蒸散面的净辐射能,LET、H分别为蒸散面与大气间的潜热和显热流交换,L为汽化潜热,G为土壤热通量。
土壤热通量与土壤中的温度梯度和热传导率成比例,
(35)
式中:
Z为深度,T为土壤温度,λ为土壤热传导率。
一般当作物温度增加时,土壤温度梯度增加,但是由于表面温度的增加通常是因为低土壤含水量引起的,当表面温度增加时,热传导率降低。
也有认为土壤热通量在数值上等于漏过冠层的净辐射,它随着叶面积指数(LAI)的增大而减小,其减小的程度大体符合比耳—朗伯定律,即:
(36)
式中:
k为消光系数。
扩散是边界层内水汽传输的
 升级会员
升级会员 