基因克隆的质粒载体Word文档下载推荐.docx
《基因克隆的质粒载体Word文档下载推荐.docx》由会员分享,可在线阅读,更多相关《基因克隆的质粒载体Word文档下载推荐.docx(22页珍藏版)》请在冰豆网上搜索。
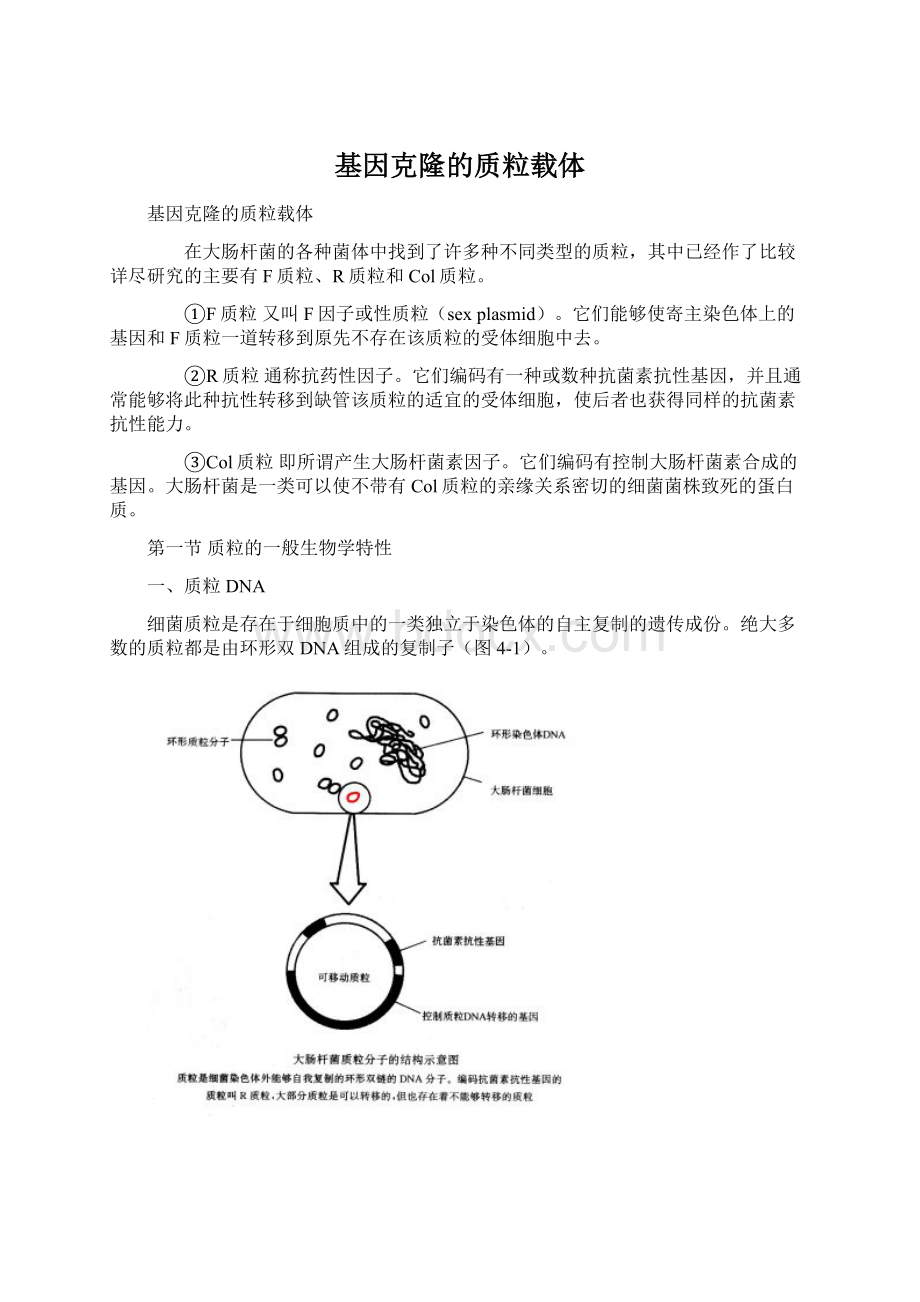
绝大多数的质粒都是由环形双DNA组成的复制子(图4-1)。
质粒DNA分子可以持续稳定地处于染色体外的游离状,但在一定的条件下又可逆地整合到寄主染色体上,随着染色体的复制而复制,并通过细胞分裂传递到后代。
环形双链的质粒DNA分子具有三种不同的构型:
1.当其两条多核苷酸链均保持着完整的环形结构时,称之为共价闭合环形DNA(cccDNA),这样的DNA通常呈现超螺旋的SC构型;
2.如果两条多核苷酸链中只有一条保持着完整的环形结构,另一条链出现有一至数个缺口时,称之为开环DNA(ocDNA),此即OC构型;
3.若质粒DNA经过适当的核酸内切限制酶切割之后,发生双链断裂形成线性分子(IDNA),通称L构型(见图4-2)。
在琼脂糖凝胶电泳中,不同构型的同一种质粒DNA,尽管分子量相同,仍具有不同的电泳迁移就绪。
其中走在最前沿的是SC DNA,其后依次是LDNA和OCDNA(图4-3)。
凡经改建而适于作为基因克隆载体的所有质粒DNA分子,都必定包括如下三种共同的组成部分,即复制基因(replicator)、选择性记和克隆位点。
二、质粒DNA编码的表型
质粒DNA仅占细胞染色体组的1%~3%左右,但却编码着一些重要的非染色体控制的遗传性状。
其中对抗菌素的抗性最质粒的最重要的编码特性之一。
三、质粒DNA的转移
(1)质粒的类型 革兰阴性细菌的质粒可以分成接合型和非接合型的两种类群。
接合型的质粒(conjugativeplasmid),又叫自我转移的质粒。
它们除了具有自主复制所必须的遗传信息之外,还带有一套控制细菌配对和质粒接合转移的基因。
非接合型的质粒(non-conjugativeplasmid),亦叫不能自我转移的质粒。
它们虽然具有自主复制的遗传信息,但失去了控制细胞配骊和接合转移的基因,因此是不能够从一个细胞自我转移到另一个细胞。
(2)F质粒 又叫F因子,即致育因子(fertilityfactor)的简称,是在某些大肠杆菌细胞中发现的一种最有代表性的单拷贝的接合型质粒。
F质粒有三种不同的存在方式:
(i)F+细胞:
以染色体外环形双链质粒DNA形式存在,其上不带有任何来自寄主染色体的基因或DNA区段。
(ii)F′细胞:
以染色体外环形双链质粒DNA形式存在,同时在其上还携带着细菌的染色体基因或DN区段。
(iii)Hfr细胞(高频重组细胞):
以线性DNA形式从不同位点整合到寄主染色体。
F因子是雄性决定因子,所以F+细胞又叫雄性细胞,与此相应的F-细胞则叫做雌性细胞。
F+细胞的表面可以形成一种叫做性须(pilus)的结构,它促进雄性细胞同雌性细胞进行配对。
在合适的条件下,将雄性细胞和雌性细胞混合培养,由于性须的作用,就会形成雌-雄细胞配对。
我们称这种过种为细菌的接合作用(conjugation)。
配对之后F-受体细胞获得了F因子,也变成为F+细胞。
由F因子整合到染色体而成的Hfr细胞,就可能相发寄主染色体发生高频转移。
这是一种可逆的过程,在一定的条件下,Hfr细胞又可重新变参展F+或F′细胞。
质粒的主要类型见表4-1。
(3)质粒DNA的接合转移
①细胞交配对的形成雄性细胞的性须顶端与受体细胞表面接触之后,便会迅速收缩,把给体细胞与受体细胞拉在一起。
因此,性须在确立配对细胞表面间的紧密接触方面,起着至关重要的作用。
但是,大肠杆菌雄性细胞是不会同其它的亦带有F质粒的细胞发生配对作用的,因为traS和traT编码的“表面排斥”蛋白质,使此种细胞无法成为接合作用的受体。
这就决定了雄性细胞只能同不具F因子的雌性细胞配对的特异性。
②质粒DNA的转移F质粒DNA的转移是从转移起点oriT开始的。
当细胞交配对建立之后,TraY和TraI蛋白质首先在oriT位点作单链切割,随后缺口链在其游离的5′-端的引导下转移到受体细胞,并作为模板合成互补链,形成新的质粒分子。
于是受体细胞便转变成为具有F因子的雄性细胞如图4-4。
四、质粒DNA的迁移作用
非接合型的质粒,由于分子小,不足以编码全部转移体系所需要的基因,因而不能够自多转移。
但如果在其寄主细胞中存在着一种接合型的质粒,那么它们通常也是可以被转移的。
这种由共存的接合型质粒引发的非接合型质粒的转移过程,叫做质粒的迁移作用(mobilization)。
ColE1是一种可以迁移但是属于非接合型的质粒。
需要质粒自己编码的两种基因参与。
一个是位于ColE1DNA上的特异位点bom;
另一个是ColE1质粒特有的弥散的基因产物,即mob基因(mobilizationgene)编码的核酸酶。
mob基因和bom基因参与ColE1质粒的迁移作用这个结论,是根据图4-5的实验结果作出的。
相容性的两种质粒F和ColE1共存于同一细菌细胞中,F质粒可以为ColE1质粒提供经所缺乏的结合功能,这样使得ColE1质粒也能够发生转移作用。
图4-5(a)表示位于F-细胞中的ColE1质粒的状,它的mob基因进行了转录,其产物使bom位点发生单链断裂而出现缺口,于是ColE1DNA便从超盘旋的的结构转变成为缺口环状的构型。
但ColE1质粒缺乏形成性须的能力,无力进行结合配对,所以它的DNA也就不能从一个细胞转移到另一个细胞。
正是由于不能够发生转移,这种从超盘旋到缺口环状的构型转变过程,就有可能被回复,所以就出现这两种构型之间的平衡状态。
图4-5(b)中的细胞同时含有F和ColE1两种质粒。
F因子能够导致性须的合成,为其DNA转移提供了转移装置,因此ColE1可以被转移。
而在F质粒提供的这种转移装置被分离掉的情况下,ColE1的mob-突变体便不能够转移。
遗传分析证明,mob-突变是隐性的,mob基因编码一种蛋白质。
而且当这种突变体质粒被分离出来时,并不是以松弛复合物的形式存在。
图4-5(c)所示,F质粒无力帮助mob-突变体进行转移,其中F性须和转移装置虽已形成,但ColE1DNA并没有发生缺口。
图4-5(d)表示另一种具mob+表型并带有一个顺式显性突变的ColE1突变体,它缺失了bom位点。
在这样的寄主细胞中,虽然能够合成mob蛋白质,但由于不能发生缺口,因此仍然不能够转移。
五、质粒DNA的复制类型
根据寄主细胞所含的拷贝数的多少,可将质粒分成两种不同的复制型:
一种是低拷贝数的质粒,每个寄主细胞中仅含有1~3份的拷贝,我们称这类质粒为“严紧型”复制控制的质粒(stringentplasmid);
另一类是高拷贝数的质粒,每个寄主细胞中可高达10~60份拷贝,这类质粒被称为“松弛型”复制控制的质粒(relaxedplasmid)。
质粒拷贝数,是指生长在标准的培养基条件下,每个细菌细胞中所含有的质粒DNA分子的数目。
表4-3列举了若干种通用的质粒复制基因的特性。
表4-2列举了若干种通用质粒复制基因的特性。
六、质粒的不亲和性
(1)质粒的不亲和性现象 所谓质粒的不亲和性(plasmidincompatibility),有时也称为不相容性,是指在没有选择压力的情况下,两种亲缘关系密切的不同质粒,不能够在同一个寄主细胞系中稳定地共存的现象。
在细胞的增殖过程中,其中必有一种会被逐渐地排斥(稀释)掉。
这样的两种质粒称为不亲和质粒(图4-6)。
不亲和群(incompatibilitygroup),指具有亲缘关系,但彼此之间是互不相容的质粒。
(2)质粒不亲和性的分子基础 质粒不亲和性的分子基础,主要是由于它们在复制功能之间的相互干扰造成的。
大多数质粒都会产生出一种控制质粒复制的阻遏蛋白质,其浓度是与质粒的拷贝数成正比的。
阻遏蛋白质通过同其靶序列间的相互作用,使双链DNA中的一条链断裂,从而导致质粒DNA复制的启动,并建立起一种调节质粒拷贝数的负反馈环(negativefeedbackloop)。
当质粒面临高拷贝数和高浓度的阻遏蛋白质时,其复制法动便被抑制了;
而当质粒处于低拷贝和低浓度遏蛋白质的条条件下,它的复制反应便会继续进行。
由于每一种质粒的复制速率拷贝数控制,都是由一对不相容质粒产生的阻遏蛋白质总浓度联合调控的,这种交叉抑制的结果,使细胞中质粒拷贝数,比其单独感染状态下的正常拷贝数减少许多。
第二节质粒DNA的复制与拷贝数的控制
一、质粒DNA复制的多样性
不同质粒DNA的复制在如下几个方面存多样性:
(i)对寄生酶的依赖性(ii)DNA聚合酶的利用(iii)复制的方向性(iv)复制的终止(v)复制型
二.ColE1质粒DNA复制的启动
ColE1质粒DNA的复制,是从一个特定的复制起点(ori)开始,并沿着环DNA分子单向性地进行。
控制此种质粒DNA复制启动的两种关键因素RNAI和RNAⅡ两种RNA分子,都是由ColE1DNA转录产生的。
其中RNAⅡ也叫做复制引物。
RNAⅡ分子在转录起点附近同互补的模板DNA形成一种杂交分子,被RnaseH酶所切割,从而释放出3′-OH末端,作为供DNA聚合酶Ⅰ合成DNA的引物。
RNAⅠ可以通过同RNAⅡ结合,以阻止其与模板DNA发生杂交作用。
因此,从本质上讲,ColE1质粒的复制启动显然是受一种负反馈机理控制的。
根据这种模型,细胞中RNAⅠ分子的浓度是随着质粒拷贝数的多寡而增减的。
例如,若细胞中质粒拷贝数下降到正常数值以下的水平,RNAⅠ的浓度也就相应降低,于是质粒的复制也就受到较少的抑制,结果导致其拷贝数的上升。
三、质粒复制控制的分子模型
目前公认的用于阐释质粒DNA复制控制机理的分子模型有两种:
其一是自体阻遏蛋白质模型(autorepressormodel),其二是抑制蛋白质稀释模型(inhibitordilutionmodel)。
前者的核心内容是,阻遏蛋白质的合成受负反馈(negativefeedback)机理调节,而且其浓度是恒定的。
后者的关键论点是,阻遏蛋白质是组成型合成,其浓度同质粒的拷贝数成正比。
(1)抑制蛋白质稀释模型
抑制蛋白质稀释模型如图4-7所示。
它认为质粒DNA的复制,是受一种由质粒DNA编码的抑制蛋白质Cop调控的。
细胞中Cop蛋白质的浓度是同质粒分子的拷贝数成正比,它抑制质粒DNA复制活性的作用方式有两种途径:
一种是通过同质粒DNA的复制起点(oir)结合,直接地抑制质粒DNA的复制;
另一种是通过阻断起始蛋白质Rep的合成,间接地抑制质粒DNA的合成。
(2)自体阻遏蛋白质模型
在抑制蛋白质稀释模型提出若干年之后,L.Sompayrac和O.Maaloe也提出了另外一种解释质粒DNA复制机理的自体阻遏蛋白质模型(图4-8)。
这个模型对质粒DNA复制控制机理有两种解释:
1.质粒DNA的复制是由一种叫做起始蛋白质Rep引发的。
Rep蛋白质编码基因rep和自体阻遏蛋白质编码基因atr是属于同一个操纵子,它们共转录成同一个mRNA分子。
自体阻遏蛋白质通过同启劝子-操纵基因区(P/O)的结合作用,调节质
 升级会员
升级会员 