第二章基因组结构与功能doc文档格式.docx
《第二章基因组结构与功能doc文档格式.docx》由会员分享,可在线阅读,更多相关《第二章基因组结构与功能doc文档格式.docx(19页珍藏版)》请在冰豆网上搜索。
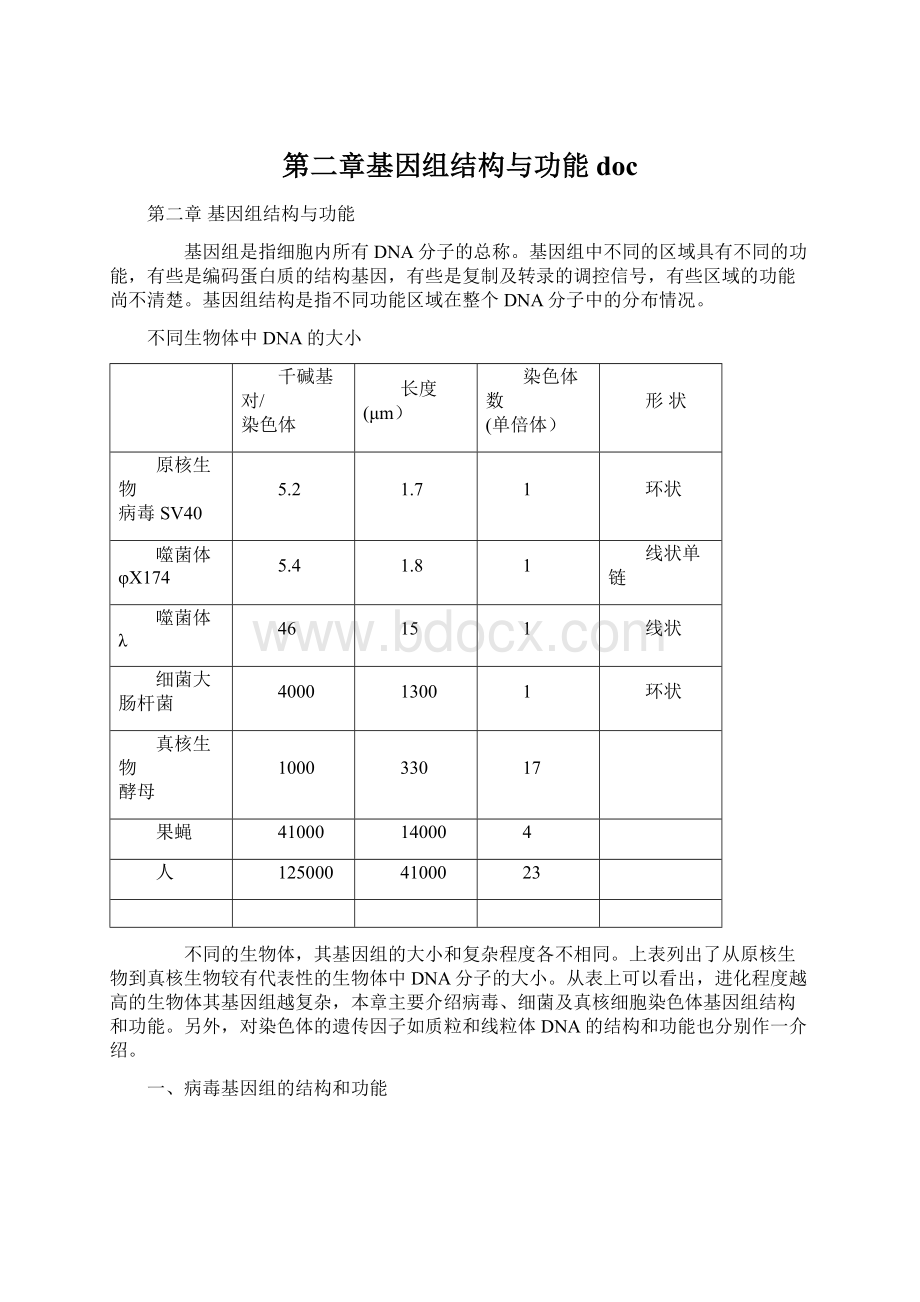
原核生物
病毒SV40
5.2
1.7
1
环状
噬菌体φX174
5.4
1.8
线状单链
噬菌体λ
46
15
线状
细菌大肠杆菌
4000
1300
真核生物
酵母
1000
330
17
果蝇
41000
14000
4
人
125000
23
不同的生物体,其基因组的大小和复杂程度各不相同。
上表列出了从原核生物到真核生物较有代表性的生物体中DNA分子的大小。
从表上可以看出,进化程度越高的生物体其基因组越复杂,本章主要介绍病毒、细菌及真核细胞染色体基因组结构和功能。
另外,对染色体的遗传因子如质粒和线粒体DNA的结构和功能也分别作一介绍。
一、病毒基因组的结构和功能
病毒是最简单的原核生物,完整的病毒颗粒包括外壳蛋白和内部的基因组DNA或RNA(有些病毒的外壳蛋白外面有一层由宿主细胞构成的被膜(envelope),被膜内含有病毒基因编码的糖蛋白。
病毒不能独立地复制,必需进入宿主细胞中借助细胞内的一些酶类和细胞器才能使病毒得以复制。
外壳蛋白(或被膜)的功能是识别和侵袭特定的宿主细胞并保护病毒基因组不受核酸酶的破坏。
1.病毒基因组的结构特点
1.与细菌或真核细胞相比,病毒的基因组很小。
如乙肝病毒DNA只有3kb大小,所含信息量也较小,只能编码4种蛋白质,而痘病毒的基因组有300kb之大,可以编码几百种蛋白质,不但为病毒复制所涉及的酶类编码,甚至为核苷酸代谢的酶类编码,因此,痘病毒对宿主的依赖性较乙肝病毒小得多。
2.病毒基因组可以由DNA或RNA组成;
病毒基因组的DNA和RNA可以是单链或双链的;
可以是闭环分子,也可以是线性分子。
如乳头瘤病毒是一种闭环的双链DNA病毒,而腺病毒的基因组则是线性的双链DNA,脊髓灰质炎病毒是一种单链的RNA病毒,而呼肠孤病毒的基因组是双链的RNA分子。
一般说来,大多数DNA病毒的基因组双链DNA分子,而大多数RNA病毒的基因组是单链RNA分子。
3.多数RNA病毒的基因组是一条RNA,但也有多条的RNA。
如流感病毒的基因组由八条RNA分子构成,每条RNA分子都含有编码蛋白质分子的信息;
而呼肠孤病毒的基因组由双链的节段性的RNA分子构成,共有10个双链RNA片段,同样每段RNA分子都编码一种蛋白质。
4.基因重叠现象普遍。
即同一段DNA片段能够编码两种甚至三种蛋白质分子,这种现象在其它的生物细胞中仅见于线粒体和质粒DNA,所以也可以认为是病毒基因组的结构特点。
这种结构使较小的基因组能够携带较多的遗传信息。
重叠基因是1977年Sanger在研究ΦX174时发现的。
ΦX174是一种单链DNA病毒,宿主为大肠杆菌,因此,又是噬菌体。
它感染大肠杆菌后共合成11个蛋白质分子,总分子量为25万左右,相当于6078个核苷酸所容纳的信息量。
而该病毒DNA本身只有5375个核苷酸,最多能编码总分子量为20万的蛋白质分子,Sanger在弄清ΦX174的11个基因中有些是重叠的之前,这样一个矛盾长时间无法解决。
重叠基因有以下几种情况:
(1)一个基因完全在另一个基因里面。
如基因A和B是两个不同基因,而B包含在基因A内。
同样,基因E在基因D内。
(2)部分重叠。
如基因K和基因A及C的一部分基因重叠。
(3)两个基因只有一个碱基重叠。
如基因D的终止密码子的最后一个碱基是J基因起始密码子的第一个碱基(如TAATG)。
这些重叠基因尽管它们的DNA大部分相同,但是由于将mRNA翻译成蛋白质时的读框不一样,产生的蛋白质分子往往并不相同。
有些重叠基因读框相同,只是起始部位不同,如SV40DNA基因组中,编码三个外壳蛋白VP1、VP2、VP3基因之间有122个碱基的重叠,但密码子的读框不一样。
而小t抗原完全在大T抗原基因里面,它们有共同的起始密码子。
5.病毒基因组的大部分是用来编码蛋白质的,只有非常小的一份不被翻译,这与真核细胞DNA的冗余现象不同如在ΦX174中不翻译的部份只占217/5375,G4DNA中占282/5577,都不到5%。
不翻译的DNA序列通常是基因表达的控制序列。
如ΦX174的H基因和A基因之间的序列(3906-3973),共67个碱基,包括RNA聚合酶结合位,转录的终止信号及核糖体结合位点等基因表达的控制区。
乳头瘤病毒是一类感染人和动物的病毒,基因组约8.0Kb,其中不翻译的部份约为1.0kb,该区同样也是其他基因表达的调控区.
6.病毒基因组DNA序列中功能上相关的蛋白质的基因或rRNA的基因往往丛集在基因组的一个或几个特定的部位,形成一个功能单位或转录单元。
它们可被一起转录成为含有多个mRNA的分子,称为多顺反子mRNA(polycistroniemRNA),然后再加工成各种蛋白质的模板mRNA。
如腺病毒晚期基因编码病毒的12种外壳蛋白,在晚期基因转录时是在一个启动子的作用下生成多顺反子mRNA,然后再加工成各种mRNA,编码病毒的各种外壳蛋白,它们在功能上都是相关的;
ΦX174基因组中的D-E-J-F-G-H基因也转录在同一mRNA中,然后再翻译成各种蛋白质,其中J、F、G及H都是编码外壳蛋白的,D蛋白与病毒的装配有关,E蛋白负责细菌的裂解,它们在功能上也是相关的。
7.除了反转录病毒以外,一切病毒基因组都是单倍体,每个基因在病毒颗粒中只出现一次。
反转录病毒基因组有两个拷贝。
8.噬菌体(细胞病毒)的基因是连续的;
而真核细胞病毒的基因是不连续的,具有内含子,除了正链RNA病毒之外,真核细胞病毒的基因都是先转录成mRNA前体,再经加工才能切除内含子成为成熟的mRNA。
更为有趣的是,有些真核病毒的内含子或其中的一部分,对某一个基因来说是内含子,而对另一个基因却是外显子。
如SV40和多瘤病毒(polyomavirus)的早期基因就是这样。
SV40的早期基因即大T和小t抗原的基因都是从5146开始反时针方向进行,大T抗原基因到2676位终止,而小t抗原到4624位即终止了,但是,从4900到4555之间一段346bp的片段是大T抗原基因的内含子,而该内含子中从4900-4624之间的DNA序列则是小t抗原的编码基因。
同样,在多瘤病毒中,大T抗原基因中的内含子则是中T和t抗原的编码基因。
2.RNA噬菌体的基因组结构和功能
目前研究最清楚的大肠杆菌RNA噬菌体是MS2,R17,f2和Qβ。
它们的基因组小,只有3600到4200个核苷酸,包含四个基因。
MS2.R17和f2具有几乎一样的基因组结构。
在四个基因中有两个基因编码噬菌体的结构蛋白:
一个是A蛋白的基因,长1178个核苷酸。
A蛋白(称为成熟蛋白)的功能是使噬菌体能识别宿主,并使其RNA基因组能进入宿主菌,每个噬菌体一般只存在分子的A蛋白。
另一个结构蛋白基因长399个核苷酸,编码外壳蛋白以构成病毒颗粒,每个噬菌体有180个分子。
基因组的其他部分编码RNA复制酶和一个溶解蛋白,编码溶解蛋白的基因与外壳蛋白和复制酶的基因有部分重叠,但读框与外壳蛋白的读框不一样。
在MS2、R17、f2基因组内有许多二级结构,RNA分子内碱基的自我配对,可能对防止RNase降解有一定作用。
另外,在编码基因的5'
和3'
端各有一段非翻译序列,该序列对稳定RNA分子也有一定作用。
另一种RNA噬菌体Qβ的基因组略大,与上述RNA噬菌体的基因组有以下不同;
[1]没有独立的溶解蛋白基因,但结构蛋白A2(或称成熟蛋白,MaturaitonProtein)即具有溶解蛋白的功能,[2]还编码另一种外壳蛋白A1。
二、细菌基因组的结构和功能
细菌基因组的结构特点在许多方面与病毒的基因组特点相似,而在另一些方面又有其独特的结构和功能。
1.细菌染色体基因组结构的一般特点
(1)组通常仅由一条环状双链DNA分子组成。
细菌的染色体相对聚集在一起,形成一个较为致密的区域,称为类核(nucleoid)。
类核无核膜与胞浆分开,类核的中央部分由RNA和支架蛋白组成,外围是双链闭环的DNA超螺旋。
染色体DNA通常与细胞膜相连。
细胞膜在这里的作用可能是对染色体起固定作用。
另外,在细胞分裂时将复制后的染色体均匀地分配到两个子代细菌中去。
有关类核结构的详细情况目前尚不清楚。
(2)具有操纵子结构。
其中的结构基因为多顺反子,即数个功能相关的结构基因串联在一起,受同一个调节区的调节。
数个操纵子还可以由一个共同的调节基因(regulatorygene)即调节子(regulon)所调控。
(3)在大多数情况下,结构基因在细菌染色体基因组中都是单拷贝,但是编码rRNA的基因rrn往往是多拷贝的,这样可能有利于核糖体的快速组装,便于在急需蛋白质合成时细胞可以在短时间内有大量核糖体生成。
(4)和病毒的基因组相似,不编码的DNA部份所占比例比真核细胞基因组少得多。
(5)具有编码同工酶的同基因(isogene)。
例如,在大肠杆菌基因组中有两个编码分支酸(chorismicacid)变位酶的基因,两个编码乙酰乳酸(acetolactate)合成酶的基因。
(6)和病毒基因组不同的是,在细菌基因组中编码序列一般不会重叠,即不出现基因重叠现象。
(7)具有各种功能的识别区域如复制起始区OriC,复制终止区TerC,转录启动区和终止区等。
这些区域往往具有特殊的序列,并且含有反向重复序列。
(8)在基因或操纵子的终末往往具有特殊的终止序列,它可使转录终止和RNA聚合酶从DNA链上脱落。
例如大肠杆菌色氨酸操纵子后尾含有40bp的GC丰富区,其后紧跟AT丰富区,这就是转录终止子的结构。
终止子有强、弱之分,强终止子含有反向重复序列,可形成茎环结构,其后面为polyT结构,这样的终止子无需终止蛋白参与即可以使转录终止。
而弱终止子尽管也有反向重复序列,但无polyT结构,需要有终止蛋白参与才能使转录终止。
2.大肠杆菌染色体基因组的结构和功能
大肠杆菌染色体基因组是研究最清楚的基因组。
估计其基因组含有3500个基因,75个操纵子。
已知基因中8%的序列具有调控作用。
大肠杆菌染色体基因组中已知的基因多是编码一些酶类的基因,如氨基酸、嘌呤、嘧啶、脂肪酸和维生素合成代谢的一些酶类的基因,以及大多数碳、氮化合物分解代谢的酶类的基因。
除了有些具有相关功能的基因在一个操纵子内由一个启动子转录外,大多数基因的相对位置可以说是随机分布的。
如控制小分子合成和分解代谢的基因,糖酵解的酶类的基因等。
在已知转录方向的50个操纵子中,27个操纵子按顺时针方向转录,23个操纵子按反时针方向转录,即DNA两条链作为模板指导mRNA合成的机率差不多相等。
在大肠杆菌染色体基因组中,差不多所有的基因都是单拷贝基因。
另外,由于大肠杆菌细胞分裂极快,可以在20分钟内完成一次分裂,因此,携带多拷贝基因的大肠杆菌并不比单拷贝基因的大肠杆菌更为有利;
相反,由于多拷贝基因的存在,使E.coli的整个基因组增大,复制时间延长,因而更为不利,除非在某种环境下,需要有多拷贝基因用来编码大量的基因产物,例如,在有极少量乳糖或乳糖衍生物的培养基上,乳糖操纵子的多拷贝化可以使大肠杆菌充分利用的乳糖分子。
但是,一旦这种选择压力消失,如将大肠杆菌移到有丰富的乳糖培养基上,多拷贝的乳糖操纵子便没有存在的必要,相反,由于需要较长的复制时间,这种重复的多拷贝基因会重新丢失。
大肠杆菌染色体基因组中,大多
 升级会员
升级会员 