医学免疫学 重点总结文档格式.docx
《医学免疫学 重点总结文档格式.docx》由会员分享,可在线阅读,更多相关《医学免疫学 重点总结文档格式.docx(18页珍藏版)》请在冰豆网上搜索。
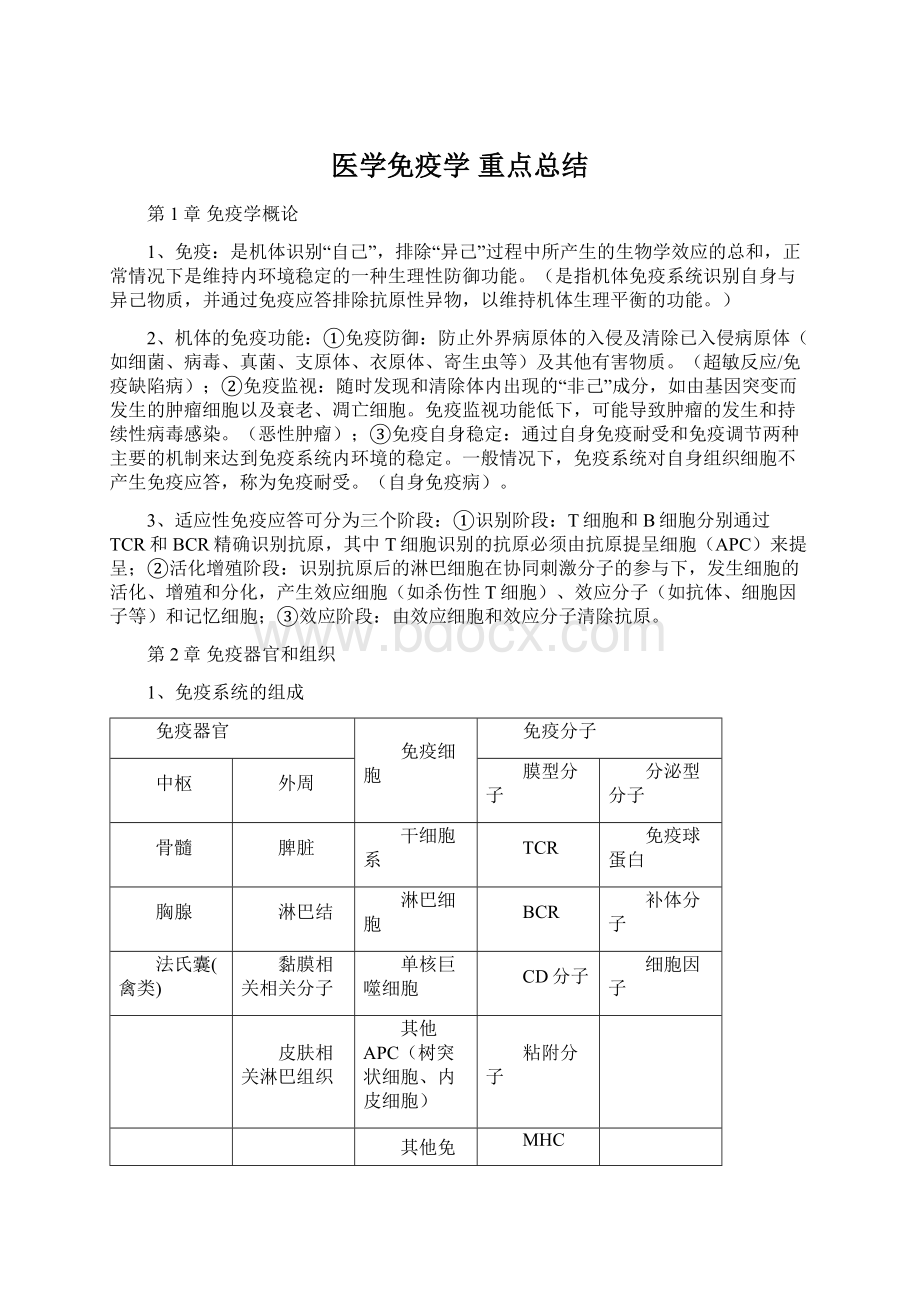
BCR
补体分子
法氏囊(禽类)
黏膜相关相关分子
单核巨噬细胞
CD分子
细胞因子
皮肤相关淋巴组织
其他APC(树突状细胞、内皮细胞)
粘附分子
其他免疫细胞(粒细胞、肥大细胞、血小板、红细胞等)
MHC
其他
2、中枢免疫器官是免疫细胞发生、分化、筛选与成熟的场所。
外周免疫器官和组织是成熟淋巴细胞(T、B细胞)定居的场所,也是这些淋巴细胞针对外来抗原刺激后启动初级免疫应答的主要部位。
3、*(n)淋巴细胞归巢:
成熟淋巴细胞离开中枢免疫器官后,经血液循环趋向性迁移并定居于外周免疫器官或组织的特定区域,称为淋巴细胞归巢。
其分子基础是淋巴细胞表面的归巢受体与内皮细胞表面相应黏附分子—血管地址素的相互作用。
4、*(n)淋巴细胞再循环:
淋巴细胞在血液、淋巴液、淋巴器官或组织间反复循环的过程,称为淋巴细胞再循环。
其生物学意义:
①使淋巴细胞在外周免疫器官和组织的分布更趋合理;
②淋巴组织可不断的得到新的淋巴细胞补充,有助于增强机体的免疫功能;
③有利于细胞识别、捕获抗原,传递免疫信息,从而产生免疫应答,增强免疫效应。
第三章抗原
1、(n)抗原(Ag):
是指能与T细胞、B细胞的TCR或BCR结合,促使其增殖、分化,产生抗体或致敏淋巴细胞,并与之结合,进而发挥免疫效应的物质。
抗原一般具备两个特性:
①免疫原性:
即抗原刺激机体产生免疫应答,诱导产生抗体或致敏淋巴细胞的能力;
②抗原性:
即抗原与其所诱导产生的抗体或致敏淋巴细胞特异性结合的能力。
2、几个概念:
(1)免疫原:
又称完全抗原,是指同时具有免疫原性和抗原性的物质。
(2)半抗原:
又称不完全抗原,即只具有抗原性而无免疫原性的物质。
(3)表位:
是与TCR/BCR及抗体特异性结合的基本单。
(4)载体:
是使半抗原变成完全抗原的物质,决定了抗原分子的免疫原性。
(半抗原若与大分子蛋白质或非抗原性的多聚赖氨酸等载体交联或结合也可以成为完全抗原。
3、异物性:
抗原免疫原性的本质是异物性。
抗原与机体之间的亲缘关系越远,组织结构差异越大,异物性越强,免疫原性就越强。
如病原微生物、血型抗原、HLA、精子、眼晶体蛋白等。
4、特异性:
抗原的特异性是指抗原刺激机体产生免疫应答及其与应答产物发生反应所显示的专一性,即某一特定抗原只能刺激机体产生特异性的抗体或致敏淋巴细胞,且仅能与该抗体或淋巴细胞发生特异性结合。
决定抗原特异性的结构基础是存在于抗原分子中的抗原表位。
(是免疫应答中最重要的特点,是免疫学诊断和免疫学防治的理论依据。
)(抗原表位:
抗原分子中决定抗原特异性的特殊化学集团,又称抗原决定簇。
是与TCR、BCR或抗体特异性结合的基本结构单位。
抗原结合价:
抗原分子上能与抗体分子结合的抗原表位的总数称为抗原结合价。
5、抗原的种类:
(1)根据诱生抗体时需否Th细胞参与分类:
①胸腺依赖性抗原(TD-Ag):
刺激B细胞产生抗体时依赖于T细胞辅助,又称T细胞依赖性抗原。
②胸腺非依赖性抗原(TI-Ag):
刺激机体产生抗体时无需T细胞的辅助,又称T细胞非依赖性抗原。
TI-Ag分为TI-1Ag和TI-2Ag,TI-1Ag具有B细胞多克隆激活作用,成熟或未成熟B细胞均可对其产生应答,TI-2Ag表面含多个重复B表位,仅能刺激成熟B细胞。
TD-Ag
TI-Ag
组成
B细胞和T细胞表位
重复B细胞表位
T细胞辅助
必需
无需
免疫应答类型
体液免疫和细胞免疫
体液免疫
抗体类型
多种
IgM
免疫记忆
有
无
(2)根据抗原与机体的亲缘关系分类:
①异嗜性抗原:
为一类与种属无关,存在于人、动物及微生物之间的共同抗原。
②异种抗原:
指来自于另一物种的抗原性物质。
如病原微生物及其产物、植物蛋白、动物免疫血清等。
③同种异型抗原:
指同一种属不同个体间所存在的抗原,又称同种抗原或同种异体抗原。
如血型抗原、组织相容性抗原。
④自身抗原:
可诱导特异性免疫应答的自身成分。
⑤独特型抗原:
TCR、BCR或Ig的V区所具有的独特的氨基酸顺序和空间构型,可诱导自体产生相应的特异性抗体,这些独特的氨基酸序列所组成的抗原表位称为独特型(Id),Id所诱生的抗体称抗独特型抗体(AId)。
(3)根据抗原是否在抗原提呈细胞内合成分类:
①内源性抗原:
在抗原提呈细胞内新合成的抗原。
如病毒蛋白、肿瘤抗原等。
②外源性抗原:
来源于抗原提呈细胞外的抗原。
如吞噬的细胞或细菌等。
(4)其他分类:
天然抗原和人工抗原,颗粒性抗原和可溶性抗原,蛋白质抗原、多糖抗原和多肽抗原,移植抗原、肿瘤抗原、变应原、过敏原及耐受原等。
6、*非特异性免疫刺激剂:
(K)
(1)超抗原(SAg):
某些物质,只需要极低浓度(1~10ng/ml)即可激活2%-20%T细胞克隆,产生极强的免疫应答,这类抗原被称为超抗原。
(2)佐剂:
预先或与抗原同时注入体内,可增强机体对该抗原的免疫应答或改变免疫应答类型的非特异性免疫增强性物质。
(3)丝裂原:
又称有丝分裂原,可导致细胞发生有丝分裂,可激活某一类淋巴细胞的全部克隆,是一种非特异性的淋巴细胞多克隆激活剂。
第四章免疫球蛋白
1、抗体(Ab):
是介导体液免疫的重要效应分子,是B细胞接受抗原刺激后增殖分化为浆细胞所产生的糖蛋白,主要存在于血清等体液中,通过与相应抗原特异性结合,发挥体液免疫功能。
2、*免疫球蛋白(Ig):
具有抗体活性或化学结构与抗体相似的球蛋白。
可分为分泌型(sIg)和膜型(mIg)。
前者主要存在于血液及组织液中,具有抗体的各种功能;
后者构成B细胞膜上的抗原受体。
3、免疫球蛋白的基本结构:
Y型结构,由两条相同的重链及两条相同的轻链,以-S-S-相连的四肽链结构。
(图P35)同一类Ig,据其铰链区氨基酸组成和重链二硫键的数目、位置不同,将其分为不同的亚类。
(1)重链(H链):
分子量50~75kDa,450-550个氨基酸残基组成。
各类免疫球蛋白重链恒定区的氨基酸组成和排列顺序不尽相同,据此可将免疫球蛋白分为5类或5个同种型:
IgA、IgG、IgM、IgD、IgE,其相应的重链分别为α链、γ链、μ链、δ链、ε链。
(2)轻链(L链):
分子量25kDa,214个氨基酸残基组成。
分κ链和λ链两种,据此将Ig分为κ型和λ型。
λ型有λ1、λ2、λ3、λ4四个亚型。
每个Ig两条L链的型别总是相同的。
两型L链的功能无差异。
不同种属生物体内两型L链的比例不同(人κ:
λ=2:
1)
(3)可变区和恒定区:
Ig轻链和重链中靠近N端氨基酸序列变化较大的区域,称为可变区(V区),分别占轻链的1/2和重链的1/4。
分VL和VH。
靠近C端氨基酸序列相对稳定的区域,称为恒定区(C区),分别占轻链的1/2和重链的3/4。
分CL和CH。
高变区(互补决定区):
VH和VL各有3个区域的氨基酸组成和排列顺序高度可变,称为高变区(HVR)或互补决定区(CDR),分别用HVR1(CDR1)、HVR2(CDR2)和HVR3(CDR3)表示,共同组成Ig的抗原结合部位,决定着抗体的特异性,负责识别及结合抗原,从而发挥免疫效应
(4)铰链区:
位于CH1与CH2之间,含有丰富的脯氨酸,因此易伸展弯曲,能改变两个结合抗原的Y形臂之间的距离,有利于两臂同时结合两个抗原表位,而且易被木瓜蛋白酶、胃蛋白酶等水解,产生不同的水解片段。
(5)结构域:
Ig分子的两条重链和两条轻链都可以折叠为数个球形结构域,每个结构与一般具有相应的功能。
Ig的二级结构是由几股多肽链折叠形成的两个反向平行的β片层,两个β片层中心的两个半胱氨酸残基由一个链内二硫键垂直连接,可稳定结构域,形成一个“β桶状”结构。
这种折叠方式称为免疫球蛋白折叠。
具有这类独特折叠结构的分子统称为免疫球蛋白超家族。
4、免疫球蛋白的其他成分:
(1)J链:
由浆细胞合成的富含半胱氨酸的多肽链,主要功能是将单体Ig分子连接为二聚体或多聚体,2个IgA单体由J链链接成二聚体,5个IgM单体通过二硫键相连,并通过二硫键与Jl链相连形成五聚体,IgG、IgD、IgE常为单体,无J链。
(2)分泌片(SP)又称额为分泌成分(SC):
由粘膜上皮细胞合成和分泌的一种含糖的肽链,是分泌型IgA(SIgA)分子上的一个辅助成分,具有保护SIgA的铰链区免受蛋白水解酶降解的作用,并介导IgA的转运。
5、免疫球蛋白的水解片段:
(1)木瓜蛋白酶水解片段:
水解部位在铰链区二硫键连接的两条H链的近N端,裂解为两个完全相同的抗原结合片段(Fab)(可与Ag结合,为单价,但不发生凝集反应或沉淀反应)和一个可结晶片段(Fc)(无抗原结合活性,是Ig与效应分子或细胞相互作用的部位)。
(2)胃蛋白酶水解片段:
水解部位在铰链区二硫键连接的两条H链的近C端,裂解为F(ab’)2(两个Fab和铰链区)(可与Ag结合,为双价,可发生凝集反应和沉淀反应)和pFc’(无生物学作用)。
6、免疫球蛋白的功能:
(1)IgV区的功能:
识别并特异性结合抗原是免疫球蛋白分子的主要功能。
①分泌型Ig:
与抗原结合后,在体内,具有中和毒素、阻断病原入侵、清除病原微生物等免疫防御功能;
在体外,有利于抗原或抗体的检测和功能的判断;
②膜型Ig:
BCR,特异性识别抗原分子。
(2)IgC区的功能:
①激活补体:
通过经典途径激活补体系统;
②结合Fc段受体:
IgG、IgA、IgE抗体可通过其Fc段与表面具有相应受体的细胞结合,产生不同的生物学作用:
1)调理作用:
指抗体如IgG的Fc段与中性粒细胞、巨噬细胞上的IgGFc受体结合,从而增强吞噬细胞的吞噬作用。
2)(n)抗体依赖的细胞介导的细胞毒作用(ADCC):
指具有杀伤活性的细胞如NK细胞通过其表面表达的Fc受体(FcR)识别包被于靶抗原(如细菌或肿瘤细胞)上抗体的Fc段,直接杀伤细胞。
3)介导Ⅰ型超敏反应。
③穿过胎盘和黏膜:
IgG是唯一能通过胎盘的免疫球蛋白,SIgA可通过呼吸道和消化道的粘膜,是粘膜局部免疫的最主要因素。
7、各类免疫球蛋白的特性与功能:
(1)IgG:
①是血清和胞外液中含量最高的Ig,约占血清总Ig的75%-80%;
②是再次免疫应答产生的主要抗体,其亲和力高,分布广泛,是机体抗感染的“主力军”;
③是唯一能通过胎盘的Ig,在新生儿抗感染免疫中起重要作用;
④可发挥激活补体、调理作用、ADCC作用等;
⑤可与葡萄球菌蛋白A结合,用于免疫诊断;
⑥某些自身抗体以及引起Ⅱ、Ⅲ型超敏反应的抗体也属于IgG。
(2)IgM:
①膜型:
单体,是BCR的主要成分,只表达mIgM是未成熟B细胞的标志;
②分泌型:
主要存在于血液中,五聚体,是分子量最大的Ig,称为巨球蛋白;
③具有很强的抗原结合能力,抗原结合价为5价;
④激活补体的能力强于IgG;
⑤天然的血型抗体为IgM;
⑥是个体发育过程中最早合成和分泌的抗体,脐带血IgM升高提示胎儿有宫内感染;
⑦是初次免疫应答中最早出现的抗体,是机体抗感染的“先头部队”,可用于感染的早期诊断。
(3)IgA:
①血清型:
单体;
二聚体,是外分泌液中的主要抗体类别,参与黏膜局部免疫,在局部抗感染中发挥重要作用,是机体抗感染的“边防军”;
③婴儿可从母亲初乳中获得SIgA,为一重要的自然被动免疫。
(4)IgD:
①分为血清型和膜结合型;
②mIgD构成BCR,是B细胞分化发育成熟的标志,未成熟B细胞仅表达mIgM,成熟B细胞同时表达mIgM和mIgD,活化的B细胞或记忆B细胞mIgD逐渐消失。
(5)IgE:
①血清中含量最少的Ig;
②亲细胞抗体,可引起Ⅰ型超敏反应;
③与机体抗寄生虫免疫有关。
第五章补体系统
存在于体液中,构成补体基本组成的蛋白质。
如:
C1~C9、MBL、B因子、D因子等
1、补体(C)系统包括30余种组分,广泛存在于血清、组织液和细胞膜表面,是一个具有精密调控机制的蛋白质反应系统。
其活化产物具有调理吞噬、溶解细胞、介导炎症、调节免疫应答和清除免疫复合物等功能。
补体固有成:
分:
2、补体系统的组成:
补体系统
补体调节蛋白:
存在于血浆中和细胞膜表面,通过调节补体激活途径中关键酶而控制补体活化强度和范围的蛋白分子。
包括H因子、I因子、S蛋白、C1抑制物等
存在于不同细胞膜表面,与补体激活过程所形成的活性片段相结合介导多种生物学效应的受体分子。
包括:
CR1-CR5、C3aR等
补体受体(CR):
3、补体激活:
其过程是一系列扩大的连锁反应,激活途径主要有:
(1)经典激活途径:
①激活物:
抗原-抗体(IgM、IgG1、IgG2、IgG3)复合物。
②激活条件:
第一个C1q分子必须同时结合两个以上IgFc段的补体结合点,才能被激活,游离的抗体不能激活补体。
(2)旁路激活途径:
某些细菌、内毒素、酵母多糖、葡聚糖。
②特点:
识别自己与非己,补体效应重要的放大机制。
(3)MBL激活途径:
病原体表面的糖结构,被甘露聚糖结合凝集素(MBL)直接识别。
②激活途径:
MBL结合细菌的甘露糖残基→结合MBL相关丝氨酸蛋白酶(MASP)→水解C4和C2……(图P50)
C4b2b3bC5-------→C5a+C5bC3bnBb↓C6、C7、C8、C9----------------→C5b6789(MAC)
4、补体活化的共同末端效应(膜攻击阶段):
三条补体活化途径形成的C5转化酶,均可裂解C5,继而通过系列的连接反应,形成C5b-C9膜攻击复合物(MAC),最终损伤靶细胞膜,致细胞崩解。
补体攻膜单位:
细胞膜表面的C3b5b与C6、C7、C8依次结合形成C5b678复和物。
该复和物诱发C9在细胞膜表面共聚,形成膜表面的通道结构MACs,造成胞膜的穿孔损伤。
5、
三条激活途径的不同点
三条激活途径的共同点:
(1)均需要激活物。
(2)均为级联反应,每一步都产生扩大效应—“滚雪球”。
(3)激活过程中均产生活性裂解片段,介导重要的生物学功能。
(4)有共同的末端效应——MAC溶解靶细胞。
(5)有某些共同的调节机制。
比较项目经典途径MBL途径旁路途径
6、
激活物抗原抗体复合物含甘露糖基的病原体表面成分病原微生物内毒素、酵母多糖等
参与成分C1~C9MBL、MASP,B、D、P因子C2~C9C3、C5~C9
抗感染作用特免效应阶段非特异性免疫非特异性免疫感染中晚期感染早期感染早期
7、补体的生物学意义:
(补体系统可参与机体的特异性与非特异性免疫应答的效应阶段,表现为生理(抗微生物防御反应、免疫调节)及病理性免疫应答(介导免疫病理的损伤性反应),是体内具有重要生物学作用的效应系统和效应放大系统)(系统性红斑狼疮CR1)
(1)补体的生物功能:
①溶菌、溶解病毒和细胞的细胞毒作用:
补体激活产生MAC,形成穿膜的亲水性通道,破坏局部磷脂双层,最终导致细胞崩解。
MAC的生物学效应是:
溶解红细胞、血小板和有核细胞;
参与宿主抗细菌和抗病毒防御机制。
②调理作用:
补体的调节吞噬作用是机体抵御全身性细菌和真菌感染的主要机制之一;
③免疫黏附作用:
机体清除循环免疫复合物的重要机制;
④炎症介质作用:
1)C3a和C5a被称为过敏毒素,它们可与肥大细胞或嗜碱性粒细胞表面C3aR和C5aR结合,触发靶细胞脱颗粒,释放组胺和其他血管活性物质,介导局部炎症反应;
2)C5a对中性粒细胞等有很强趋化活性,可诱导中性粒细胞表达黏附分子,刺激中性粒细胞产生氧自由基、前列腺素和花生四烯酸,引起血管扩张、毛细血管通透性增高、平滑肌收缩等。
(2)补体的病理生理学意义:
(1)机体抗感染防御的主要机制:
①旁路途经是最早出现的C3活化途径;
②MBL途径将原始的、凝集素介导的防御功能与补体相联系,更显示补体作为固有免疫防御机制的重要性;
③补体经典途径在种系发生上出现最晚,它将非特异的补体与特异的适应性免疫相联系,成为体液免疫应答的重要效应机制。
(2)参与适应性免疫应答(免疫应答的诱导/免疫细胞增殖分化/免疫应答效应/免疫记忆)。
(3)与血液中其他级联反应系统相互作用(补体与凝血系统、激肽系统和纤溶系统等存在相互作用)。
第六章细胞因子
1、细胞因子:
是由免疫原、丝裂原或其他因子刺激细胞所产生的低分子量可溶性蛋白质,为生物信息分子,具有调节固有免疫和适应性免疫应答,促进造血,以及刺激细胞活化、增殖和分化等功能。
2、*细胞因子的产生细胞:
淋巴细胞、单核-巨噬细胞、树突状细胞、粒细胞以及其他非造血细胞(如成纤维细胞和内皮细胞等)等;
辅助性T淋巴细胞(Th)是产生细胞因子最多的免疫细胞。
3、细胞因子的共同特点:
①多为小分子(8~30kD)多肽;
②在较低浓度下即有生物学活性;
③通过结合细胞表面高亲和力受体发挥生物学效应;
④以自分泌、旁分泌或内分泌形式发挥作用;
⑤具有多效性、重叠性、拮抗性或协同性。
5、细胞因子的生物学活性:
*①调节固有免疫应答;
②调节适应性免疫应答;
③刺激造血;
④促进凋亡,直接杀伤靶细胞;
⑤促进创伤的修复。
6、细胞因子受体:
*结构:
均为跨膜蛋白,由胞膜外区、跨膜区和胞质区组成。
分类:
免疫球蛋白超家族受体、Ⅰ类细胞因子受体、Ⅱ类细胞因子受体、肿瘤坏死因子受体超家族和趋化细胞因子受体等。
第8章主要组织相容性复合体及其编码分子
1、*主要组织相容性复合体(MHC):
是脊椎动物某一染色体上编码主要组织相容性抗原、控制细胞间相互识别、将抗原信息提呈给CD4+的T细胞、调节免疫应答的一组紧密连锁的基因群。
在人类MHC称为HLA基因或HLA复合体,其产物为HLA分子或HLA抗原。
目前已经证实MHC不仅与移植有关,而且也与免疫应答和调节有关。
2、*组成MHC的各种基因传统上分为I类、II类和III类,晚近倾向于以两种类型加以概括:
一是经典的I类基因和II类基因,它们的产物具有抗原提呈功能,显示极为丰富的多态性,直接参与T细胞的激活和分化,参与调控适应性免疫应答。
二是免疫功能相关基因,包括传统的III类基因,以及新近确认的多种基因,它们主要参与调控固有免疫应答,不显示或显示有限的多样性。
3、HLA与临床医学:
器官移植的成败主要取决于供、受者间的组织相容性,其中HLA等位基因的匹配程度尤为重要。
强直性脊柱炎:
HLA-B27抗原阳性率高达58%~97%。
4、经典的MHCI类基因和II类基因:
(1)定位于小鼠第17号染色体的H-2,长约1500kb。
其中I类基因包括K、D、L三个座位,分布于复合体两侧。
II类基因由四个座位组成,称为Ab、Aa、Eb和Ea,分别编码Aβ、Aα、Eβ和Eα四种肽链。
肽链Aβ和Aα形成异二聚体,称I-A分子;
Eβ和Eα形成异二聚体,成I-E分子。
在A和E前面均加字母I,代表其编码基因坐落在H-2复合体的免疫应答区即I区。
(2)HLA基因复合体位于人第6染色体短臂上,全长3600kb,224个基因座位,128个功能基因。
经典的HLAI类基因集中在远离着丝点的一端,按序包括B、C、A三个座位,产物称作HLAI类分子。
I类基因仅编码I类分子异二聚体中的重链,轻链为β2微球蛋白(β2-m),其编码基因位于第15号染色体。
HLAII类分子由HLAII类基因编码,HLAII类基因在复合体中靠近着丝点,结构较为复杂,顺序由DP、DQ、和DR三个亚区组成。
每一亚区又包括了两个或以上的功能基因作为,分别编码分子量相近的α链和β链,形成DRα-DRβ、DQα-DQβ、DPα-DPβ三种异二聚体。
(*HLAI类和II类等位基因产物(HLA分子)的表达具有共显性特点,即同源染色体对应座位上的两个等位基因皆能得到表达,因而一个免疫细胞的表面通常可以检测到分别来自父母双方六对共12种HLAI类和II类等位基因分子即HLA抗原。
①HLA-Ⅰ类基因:
包括A、B、C三个座位,编码HLA-I类分子的重链。
②HLA-Ⅱ类基因:
包括DP、DQ、DR三个亚区,编码HLA-Ⅱ类分子。
③HLA-Ⅲ类基因:
编码补体分子。
)(图P77)
HLA抗原类别
座位
分子结构
肽结合结构域
表达特点
组织分布
功能
I类
A、B、C
α链45kDβ2-m12kD
α1+α2
共显性
所有有核细胞表面
识别和提呈内源性抗原肽,与辅助受体CD8结合,对CTL的识别起限制作用
II类
DR、DQ、DP
α链35kDβ链28kD
α1+β1
APC、火化的T细胞
识别和提呈外源性抗原肽,与辅助受体CD4结合,对Th的识别起限制作用
第9章B淋巴细胞
1、*B淋巴细胞,由哺乳动物骨髓或鸟类法氏囊中的淋巴样干细胞分化发育而来。
成熟B细胞主要定居于外周淋巴器官的淋巴小结内。
B细胞不仅能通过产生抗体发挥特异性体液免疫功能,也是重要的抗原提呈细胞。
B细胞在中枢免疫器官中的分化发育过程中发生的主要事件是功能性B细胞(抗原)受体(BCR)的表达和自身免疫耐受的形成。
2、等位排斥:
B细胞只表达一条染色体上的轻链或重链基因,先表达基因将抑制另一条染色体上的基因。
同种型排斥:
一个克隆的B细胞只能表达一种轻链。
即在合成抗原识别受体时只表达一种轻链,即两种轻链的表达存在排斥。
3、*抗原识别受体多样性产生的机制:
(1)组合造成的多样性:
抗原识别受体的胚系基因以众多的V、D、J片段成簇存在,当发生V、(D)、J基因片段重排时,只能分别取用众多V、(D)、J基因片段中的1个,因而可以产生众多V区基因片段组合。
(2)连接造成的多样性:
Ig各基因片段之间的连接往往并不准确,有插入、替换或缺失核苷酸的情况发生,从而产生新的序列,称为连接多样性。
(3)体细胞高频率突变造成的多样性:
是在已成熟B细胞即已完成V基因重排的基础上发生的,而且发生在抗原刺激后外周淋巴器官生发中心的B细胞,主要是发生在V区的CDR区上的点突变,还可导致抗体的亲和力成熟。
4、*B细胞在中枢免疫器官中的分化发育:
(1)祖B细胞:
早期祖细胞,其重链可变区基因D-J开始重排;
(2)前B细胞:
经历大前B细胞和小前B细胞(其轻链基因发生V-J重排)两个阶段,前B细胞受体由μ链和替代轻链(λ5和VpreB)所组成;
(3)为成熟B细胞:
可以表达完整mlgM;
(4)成熟B细胞:
可以同时表达mlgM、mlgD,离开骨髓。
5、*B细胞中枢免疫耐受的形成:
①克隆清除:
未成熟B细胞
 升级会员
升级会员 